Сравнительная характеристика конкурентного и неконкурентного ингибирования. Ингибиторы ферментов. Регуляция по типу обратной связи
Читайте также
Различают обратимое и необратимое ингибирование. Если ингибитор вызывает стойкие изменения пространственной третичной структуры молекулы фермента или модификацию функциональных групп фермента, то такой тип ингибирования называется необратимым. Чаще, однако, имеет место обратимое ингибирование, поддающееся количественному изучению на основе уравнения Михаэлиса-Ментен. Обратимое ингибирование в свою очередь разделяют на конкурентное и неконкурентное в зависимости от того, удается или не удается преодолеть торможение ферментативной реакции путем увеличения концентрации субстрата.
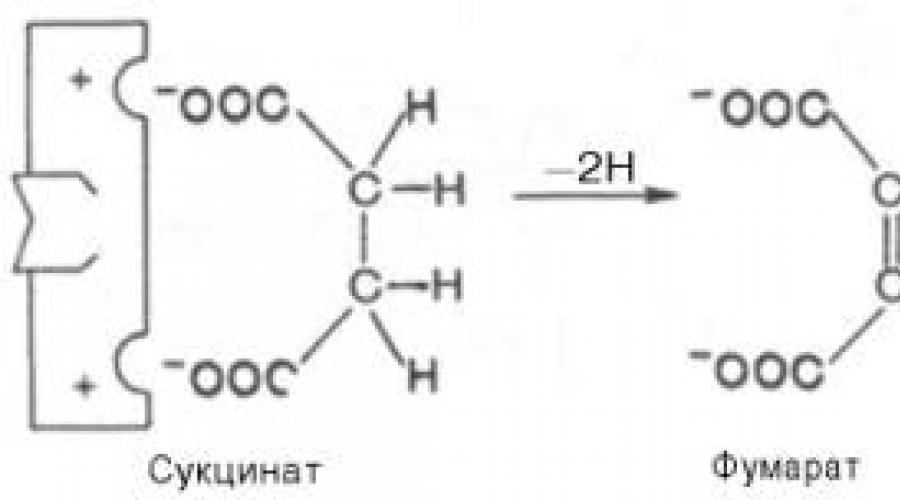
Конкурентное ингибирование может быть вызвано веществами, имеющими структуру, похожую на структурусубстрата, но несколько отличающуюся от структуры истинного субстрата. Такое ингибирование основано на связывании ингибитора с субстратсвязывающим (активным) центром. Классическим примером подобного типа ингибирования является торможение сукцинатдегидрогеназы (СДГ) малоновой кислотой. Этот ферменткатализирует окисление путем дегидрирования янтарной кислоты (сукцината) в фумаровую:
Если в среду добавить малонат (ингибитор), то в результате структурного сходства его с истиннымсубстратом сукцинатом (наличие двух таких же ионизированных карбоксильных групп) он будет взаимодействовать с активным центром с образованием фермент-ингибиторного комплекса, однако при этом полностью исключается перенос атома водорода от малоната. Структуры субстрата (сукцинат) и ингибитора(малонат) все же несколько различаются. Поэтому они конкурируют за связывание с активным центром, и степень торможения будет определяться соотношением концентраций малоната и сукцината, а не абсолютной концентрацией ингибитора. Таким образом, ингибитор может обратимо связываться сферментом, образуя фермент-ингибиторный комплекс. Этот тип ингиби-рования иногда называют ингибированием по типу метаболического антагонизма (рис. 4.20).
В общей форме реакция взаимодействия ингибитора с ферментом может быть представлена следующим уравнением:
Образовавшийся комплекс, называемый фермент-ингибиторным комплексом ЕI, в отличие от фермент-субстратного комплекса ES не распадается с образованием продуктов реакции. Константу диссоциациикомплекса EI, или ингибиторную константу К i , можно, следуя теории Михаэлиса–Мен-тен, определить как отношение констант обратной и прямой реакций:
Метод конкурентного торможения нашел широкое применение в медицинской практике. Известно, например, что для лечения некоторых инфекционных заболеваний, вызываемых бактериями, применяютсульфаниламидные препараты. Оказалось, что эти препараты имеют структурное сходство спарааминобензойной кислотой, которую бактериальная клетка использует для синтеза фолиевой кислоты, являющейся составной частью
Рис. 4.20. Действие конкурентного ингибитора (схема по В.Л. Кретовичу). Е - фермент; S - субстрат; Р 1 и Р 2 - продукты реакции; I - ингибитор.
ферментов бактерий. Благодаря этому структурному сходству сульфаниламид блокирует действие ферментапутем вытеснения парааминобензой-ной кислоты из комплекса с ферментом, синтезирующим фолиевую кислоту, что ведет к торможению роста бактерий.
Неконкурентное ингибирование вызывается веществами, не имеющими структурного сходства с субстратами и часто связывающимися не с активным центром, а в другом месте молекулы фермента. Степень торможения во многих случаях определяется продолжительностью действия ингибитора на фермент. При данном типе ингибирования благодаря образованию стабильной ковалентной связи фермент часто подвергается полной инактивации, и тогда торможение становится необратимым. Примером необратимого ингибирования является действие йодацетата, ДФФ, а также диэтил-n-нитрофенилфосфата и солей синильной кислоты. Это действие заключается в связывании и выключении функциональных групп или ионов металлов и молекулефермента.
Оглавление темы "Ингибирование. Клетка.":
В этом случае вещество, близкое по своей структуре к обычному субстрату фермента , соединяется с активным центром фермента, но не может прореагировать с ним. Находясь здесь, оно преграждает доступ к активному центру любой молекуле настоящего субстрата.
Поскольку в этом случае ингибитор и субстрат конкурируют за место на активном центре фермента , эту форму ингибирования называют конкурентным ингиЖированием. Для конкурентного ингибирования характерно, что, если концентрация субстрата увеличивается, то скорость реакции возрастает, т. е. это ингибирование обратимо.
Рисунок иллюстрирует один из примеров конкурентного ингибирования .
Явление конкурентного ингибирования помогает понять механизм действия некоторых лекарственных препаратов, в частности сульфаниламидов. Цель химиотерапии - уничтожить при помощи тех или иных химических препаратов возбудителя болезни, не повреждая при этом ткани организма-хозяина. Первыми такими препаратами были сульфаниламиды, антибактериальное действие которых было обнаружено в 30-е годы XX в. Во время второй мировой войны их широко применяли для борьбы с раневыми инфекциями. Сульфаниламиды по своей химической природе близки к парааминобензойной кислоте (ПАБК) - необходимому фактору роста многих патогенных бактерий. ПАБК требуется бактериям для синтеза фолиевой кислоты, которая служит у них одним из кофакторов ферментов. Сульфаниламиды ингибируют один из ферментов, участвующих в синтезе фолиевой кислоты из ПАБК.
Животные клетки нечувствительны к сульфаниламидам , хотя им для некоторых реакций и требуется фолиевая . Объясняется это тем, что они используют уже образованную фолиевую кислоту; метаболический путь, который бы обеспечивал ее синтез, у них отсутствует.
Неконкурентное обратимое ингибирование
Ингибиторы этого типа не родственны по своей структуре субстрату данного фермента; в образовании комплекса с ингибитором участвует в этом случае не активный центр фермента, а какая-нибудь другая часть его молекулы. Это не препятствует соединению субстрата с ферментом, но делает невозможным катализ.
С повышением концентрации ингибитора скорость реакции все более снижается. К моменту насыщения ингибитором она оказывается практически равной нулю. В отличие от конкурентного инги-бирования в этом случае повышение концентрации субстрата на скорость реакции не влияет.
Ингибирование
– это торможение активности фермента. При этом денатурации ферментов не происходит.
Ингибитор - вещество, вызывающее специфичное снижение активности фермента. Неорганические кислоты и тяжелые металлы ингибиторами не являются, а являются инактиваторами , так как снижают активность любых ферментов, т.е. действуют неспецифично .Так же денатурирующие агенты к ингибитррам не относят.
Ингибиторы: ионы или небольшие молекулы, составляющие часть ферментативной регуляторной системы, а так же фармакологические препараты.
по прочности связывания фермента с ингибитором ингибирование бывает обратимым и необратимым .
по отношению ингибитора к активному центру фермента ингибирование делят на конкурентное и неконкурентное .
Виды ингибирования
1. Обратимое 2. Необратимое
А. КОНКУРЕНТНОЕ А. СПЕЦИФИЧЕСКОЕ
Б. НЕКОНКУРЕНТНОЕ Б. НЕСПЕЦИФИЧЕСКОЕ
Обратимое ингибирование . Большинство ингибиторов действуют обратимо, образуя нековалентные связи с ферментом , и при определенных условиях диссоциируют с восстановлением активности фермента.
Конкурентное ингибирование. Ингибитор похож на субстрат фермента по своей структуре и соперничает с субстратом за активный центр (садится на активный центр фермента ), что приводит к уменьшению связывания субстрата с ферментом и нарушению катализа. В этом состоит особенность конкурентного ингибирования – возможность усилить или ослабить ингибирование через изменение концентрации субстрата.
Для конкурентного типа ингибирования справедливы следующие уравнения:
Е + S ⇔ ES → E + P,
1. Конкурентное взаимодействие этанола и метанола за активный центр алкогольдегидрогеназы .
2. Ингибирование сукцинатдегидрогеназы малоновой кислотой , структура которой схожа со структурой субстрата этого фермента – янтарной кислоты (сукцината).

Сукцинат + ФАД ----------- Фумарат + ФАДН 2
3.Также к конкурентным ингибиторам относят антиметаболиты или псевдосубстраты, например, антибактериальные средства сульфаниламиды , схожие по структуре с п -аминобензойной кислотой, компонентом фолиевой кислоты. При лечении сульфаниламидами в бактериальной клетке конкурентно нарушается использование п -аминобензойной кислоты для синтеза фолиевой кислоты , что и вызывает лечебный эффект.

Сходство строения сульфаниламидов и парааминобензойной кислоты, компонента витамина в9

Влияние различных концентраций субстрата на скорость реакции, катализируемой ферментами 1 и 2(в присутствии ингибитора) : а) гиперболическая зависимость V от [ S ], б) прямая зависимость в обратных координатах 1/ V от 1/[ S ] - Лайнуивера-Бэрка.
Конкурентные ингибиторы уменьшают скорость химической реакции. Конкурентный ингибитор повышает К m для данного субстрата (уменьшает сродство субстрата к ферменту). Это означает, что в присутствии конкурентного ингибитора необходима большая концентрация субстрата для достижения 1/2 V max . Увеличение соотношения концентрации субстрата и ингибитора снижает степень ингибирования. При значительно более высоких концентрациях субстрата ингибирование полностью исчезает , потому что активные центры всех молекул фермента будут находиться преимущественно в комплексе с субстратом.
Неконкурентное ингибирование. Ингибитор не имеет структурного сходства с субстратом и присоединеняется не в активном центре , а в другом месте молекулы, одновременно с субстратом. Образуется тройной комплекс: субстрат - фермент - ингибитор. Это ведет к деформации активного центра и каталитической активности. Например, синильная кислота (цианиды) связывается с гемовым железом ферментов дыхательной цепи и блокирует клеточное дыхание.

Кинетическая зависимость неконкурентного ингибирования: характеризуется снижением V max ферментативной реакции и уменьшением сродства субстрата к ферменту, т.е. увеличением К m .

Неконкурентное ингибирование в двойных обратных координатах при различных концентрациях ингибитора (1 - [I]=0; 2 - [I]>0; 3 - [I]>[I]2).
При неконкурентном ингибировании константа Михаэлиса не изменяется, а максимальная скорость реакции уменьшается в (1 + [I ]/K i ) раз. Поэтому в двойных обратных координатах семейство прямых, отвечающих разным концентрациям ингибитора, пересекается в одной точке на оси абсцисс.Необратимое ингибирование наблюдают в случае образования ковалентных стабильных связей между молекулой ингибитора и фермента. Чаще всего модификации подвергается активный центр фермента, В результате фермент не может выполнять каталитическую функцию.
К необратимым ингибиторам относят ионы тяжёлых металлов, например ртути (Hg 2+), серебра (Ag +) и мышьяка (As 3+), которые в малых концентрациях блокируют сульфгидрильные группы активного центра. Субстрат при этом не может подвергаться химическому превращению (рис. 2-26). При наличии реактиваторов ферментативная функция восстанавливается. В больших концентрациях ионы тяжёлых металлов вызывают денатурацию белковой молекулы фермента, т.е. приводят к полной инактивации фермента.
Оренбург – 2010
1.1 Обратимое ингибирование
1.1.2 Неконкурентное ингибирование
1.1.3 Бесконкурентное ингибирование
1.2 Необратимое ингибирование
1.3 Аллостерическое ингибирование
2. Новый вид ингибирования ферментативной активности
3. Применение ингибиторов ферментов
ЗАКЛЮЧЕНИЕ
Список использованной литературы
1. Ингибиторы ферментов. Типы ингибирования активности ферментов
Известно, что активность ферментов сравнительно легко может быть уменьшена с помощью разнообразных воздействий. Такое снижение скорости ферментативных реакций принято называть торможением активности, или ингибированием ферментов.
Рис 1. Схема активирования и ингибирования действия фермента (по Ю. Б. Филипповичу): а. – аллостерический центр фермента; К - каталитический центр; с - субстратный центр
Ферменты являются белками, соответственно их активность можно снизить или полностью ликвидировать путем воздействий, приводящих к денатурации белков (нагревание, действие концентрированных кислот, щелочей, солей тяжелых металлов и т.п.) Это неспецифическое подавление активности ферментов, имеющее значение при изучении ферментативных реакций, не представляет особого интереса для исследования их механизма. Гораздо большее значение имеет исследование ингибирования с помощью веществ, специфически и обычно в небольших количествах взаимодействующих с ферментами – ингибиторов ферментов. Расшифровка механизмов многих биологических процессов, таких как гликолиз, цикл Кребса и других, стала возможной лишь в результате применения специфических ингибиторов различных ферментов (Н.Е. Кучеренко, Ю.Д. Бабенюк и др., 1988).
Некоторые ингибиторы ферментов являются для организма животных и человека эффективными лекарственными веществами, другие - смертельными ядами (В.П. Комов, В.Н. Шведова, 2004).
Ингибиторы взаимодействуют с активными центрами молекулы фермента, инактивируя функциональные группы белков. Они могут взаимодействовать с металлами, входящими в состав молекул ферментов и фермент-субстратных комплексов, инактивируя их. Высокие концентрации ингибиторов разрушают четвертичную, третичную и вторичную структуры молекулы фермента, вызывая его денатурацию (А.И. Кононский, 1992).
Недавно открыты антиферменты (антиэнзимы, или антизимы), представляющие собой белки, действующие как ингибиторы ферментов. К подобным веществам относятся, например, ингибитор трипсина, обнаруженный в соевых бобах, и сывороточный антитрипсин. Недавно открыт в печени животных антифермент орнитиндекарбоксилазы. Антизимы, вероятнее всего, образуют труднодиссоциируемые комплексы с соответствующими ферментами, выключая их из химических реакций. Иногда ингибитор является составным компонентом предшественника фермента, или входит в состав сложных комплексов ферментов. Однако до сих пор не выяснено, являются ли подобные антиферменты истинными ингибиторами или регуляторными субъединицами.
Если ингибитор вызывает стойкие изменения пространственной третичной структуры молекулы фермента или модификацию функциональных групп фермента, то такой тип ингибирования называется необратимым. Чаще, однако, имеет место обратимое ингибирование, поддающееся количественному изучению на основе уравнения Михаэлиса-Ментен. Обратимое ингибирование в свою очередь разделяют на конкурентное и неконкурентное
На практике многие ингибиторы не проявляют тех свойств, которые характерны для чисто конкурентного или чисто неконкурентного ингибирования. Другой способ классификации ингибиторов основывается на характере места их связывания. Одни из них связываются с ферментом в том же месте, что и субстрат (в каталитическом центре), а другие - на значительном расстоянии от активного центра (в аллостерическом центре) (Р. Марри, Д. Греннер и др., 1993).
1.1 Обратимое ингибирование
Различают три типа обратимого ингибирования ферментов: конкурентное, неконкурентное и бесконкурентное, в зависимости от того, удается или не удается преодолеть торможение ферментативной реакции путем увеличения концентрации субстрата.
1.1.1 Конкурентное ингибирование
Конкурентный ингибитор конкурирует с субстратом за связывание с активным центром, но в отличие от субстрата связанный с ферментом конкурентный ингибитор не подвергается ферментативному превращению. Отличительная особенность конкурентного ингибирования состоит в том, что его можно устранить или ослабить, просто повысив концентрацию субстрата. Например, если при заданных концентрациях субстрата и конкурентного ингибитора активность фермента подавлена на 50 %, то мы можем уменьшить степень ингибирования, повысив концентрацию субстрата.
По своей трехмерной структуре конкурентные ингибиторы обычно напоминают субстрат данного фермента. Благодаря такому сходству конкурентному ингибитору удается "обмануть" фермент и связаться с ним. Конкурентное ингибирование можно количественно изучать на основе теории Михаэлиса-Ментен. Конкурентный ингибитор I просто обратимо присоединяется к ферменту Е, образуя с ним комплекс
Конкурентное ингибирование проще всего можно распознать экспериментальным путем, определив влияние концентрации ингибитора на зависимость начальной скорости реакции от концентрации субстрата. Для выяснения вопроса о том, по какому типу - конкурентному или неконкурентному - происходит обратимое ингибирование фермента, используют метод двойных обратных величин. Из графиков, построенных в двойных обратных координатах, можно определить также значение константы диссоциации комплекса фермент ингибитор (см. рис. 1) (А. Ленинджер, 1985)
Конкурентное ингибирование может быть вызвано веществами, имеющими структуру, похожую на структуру субстрата, но несколько отличающуюся от структуры истинного субстрата. Такое ингибирование основано на связывании ингибитора с субстратсвязывающим (активным) центром (см. рис. 2).

Рис. 2. Общий принцип конкурентного ингибирования (схема по В.Л. Кретовичу). Е - фермент; S - субстрат; Р 1 и Р 2 - продукты реакции; I - ингибитор.
В качестве примера можно привести действие малоновой кислоты на реакцию, которая катализируется сукцинатдегидрогеназой и связана с превращением янтарной кислоты в фумаровую. Добавление малоновой кислоты к реакционной смеси снижает или полностью останавливает ферментативную реакцию, так как она является конкурентным ингибитором сукцинатдегидрогеназы. Сходства малоновой кислоты с янтарной достаточно для образования комплекса с ферментом, однако распад этого комплекса не происходит. При увеличении концентрации янтарной кислоты она вытесняет малоновую кислоту из комплекса, в результате активность сукцинатдегидрогеназы восстанавливается.

Рис. 3. Конкурентное ингибирование реакции превращения янтарной кислоты в фумаровую под действием малоновой кислоты.
Структуры субстрата (сукцинат) и ингибитора (малонат) все же несколько различаются. Поэтому они конкурируют за связывание с активным центром, и степень торможения будет определяться соотношением концентраций малоната и сукцината, а не абсолютной концентрацией ингибитора. Таким образом, ингибитор может обратимо связываться с ферментом, образуя фермент-ингибиторный комплекс. Этот тип ингибирования иногда называют ингибированием по типу метаболического антагонизма (см. рис. 3).
В общей форме реакция взаимодействия ингибитора с ферментом может быть представлена следующим уравнением:

Образовавшийся комплекс, называемый фермент-ингибиторным комплексом ЕI, в отличие от фермент-субстратного комплекса ES не распадается с образованием продуктов реакции.
Многие лекарственные вещества ингибируют ферменты человека и животных по конкурентному типу. Например, что для лечения некоторых инфекционных заболеваний, вызываемых бактериями, применяют сульфаниламидные препараты. Оказалось, что эти препараты имеют структурное сходство с парааминобензойной кислотой, которую бактериальная клетка использует для синтеза фолиевой кислоты, являющейся составной частью ферментов бактерий. Благодаря этому структурному сходству сульфаниламид блокирует действие фермента путем вытеснения парааминобензойной кислоты из комплекса с ферментом, синтезирующим фолиевую кислоту, что ведет к торможению роста бактерий.

В структуру пептидогликана клеточной стенки бактерий включен D-аланин, отсутствующий в организме животных и человека. Для синтеза клеточной стенки бактерии при помощи фермента аланин-рацемазы превращают L-аланин животных в D-форму. Аланин-рацемаза характерна для бактерий и не обнаружена у млекопитающих. Следовательно, она представляет хорошую мишень для ингибирования лекарственными препаратами. Замещение одного из протонов метильной группы на фтор дает фтораланин, с которым связывается аланин-рацемаза, что приводит к ее ингибированию.