Кто ввел в науку термин ген. Ген — Медицинская энциклопедия. Изменения хромосом в процессе деления клетки
Читайте также
Понятие "ген" возникло задолго до возникновения науки, его изучающей. Чешский естествоиспытатель, основатель современной генетики, Гргеор Мендель в 1865 г. анализируя опыты по скрещиванию гороха, пришел к выводу, что наследование признаков осуществляется дискретными частицами, которые он называл "зачатками" или наследственными "факторами". В 1868 году Чарльз Дарвин предложил "временную гипотезу" пангенеза, согласно которой все клетки организма отделяют от себя особые частицы, или геммулы, а из них, в свою очередь, образуются половые клетки.
Затем Гуго де Фриз в 1889 году, спустя 20 лет после Ч. Дарвина, выдвинул свою гипотезу внутриклеточного пангенеза и ввел термин "панген" для обозначения имеющихся в клетках материальных частиц, которые отвечают за вполне конкретные отдельные наследственные свойства, характерные для данного вида. Геммулы Ч. Дарвина представляли ткани и органы, пангены де Фриза соответствовали наследственным признакам внутри вида.
В 1906 г. английским ученым У. Бетсоном было введено название науки - "генетика", а спустя три года, в 1909 г. , датский ученый В. Иогансен счёл удобным пользоваться только второй частью термина Гуго де Фриза "ген" и заменить им неопределенное понятие "зачатка", "детерминанта", "наследственного фактора". При этом В. Иогансен подчеркивал, что "этот термин совершенно не связан ни с какими гипотезами и имеет преимущество вследствие своей краткости и легкости, с которой его можно комбинировать с другими обозначениями". Он сразу же образовал ключевое производное понятие "генотип" для обозначения наследственной конституции гамет и зигот в противоположность фенотипу. Таким образом в генетику вошло понятие гена как элементарной единицы наследственности. В дальнейшем оно постоянно уточнялось благодаря многочисленным открытиям: была доказана локализация генов в хромосомах; выяснилось, что гены изменяются в результате мутаций; было разработано понятие об аллелях и их локализации в соответствующих локусах гомологичных хромосом. Во всех генетических исследованиях ген становится общепризнанной единицей наследственности.
Среди генетиков было всеобщим убеждение в неделимости гена. Они представляли себе ген как единое целое, как последнюю элементарную единицу наследственности. Но уже в начале 30-х годов возникло сомнение в том, что ген неделим. Первым сигналом в этом отношении положило открытие множественных аллелей, или серии множественных аллелей. Получилось так, что единый ген может изменяться, давая целый ряд мутаций, связанных с изменениями определенного признака.
У некоторых организмов, и прежде всего у дрозофилы, были открыты серии множественных аллелей, содержащих десятки разнообразных мутаций, а у рогатого скота обнаружена серия аллелей, включающая до 80 мутаций, т. е. в результате мутаций возникло 80 разных состояний одного локуса.
С начала 30-х годов начался новый этап в изучении гена. Разработкой его строения была занята лаборатория А. С. Серебровского. Работы А. С. Серебровского, затем Н. П. Дубинина показали, что ген имеет значительно более сложное строение, чем предполагали раньше.
Работы велись по изучению гена scute, локализованного в половой хромосоме дрозофилы. Этот ген определяет развитие щетинок на теле мухи. Различные аллельные мутации гена касались недоразвития щетинок на тех или иных определенных участках тела дрозофилы и различной степени редукции щетинок. При генетическом анализе этих мутаций, скрещивании их друг с другом выяснилось, что в гетерозиготе они ведут себя частично как аллельные гены, а частично как мутации независимых локусов хромосом. Таким образом, ген оказался сложной системой, в которой мутации ведут к изменению лишь отдельных его частей.
Название "множественные аллели" было заменено более удачным "ступенчатые аллели" и была сформулирована гипотеза о сложном строении гена. Ген в целом назван "базигеном", а мутировавшие аллели "трансгенами".
Дальнейшее развитие учения о строении гена связано с переходом методов генетических исследований с хромосомного на молекулярный уровень. Большое значение при этом имело использование в работах генетиков до того времени мало изученных микроорганизмов: бактерий и даже неклеточных форм - вирусов. Особенно большое значение в этих работах имели исследования бактериофагов из группы "Т", заражающих кишечную палочку.
В изучении природы гена особенно большое значение имели работы Бензера и ряда других исследователей, проведенные на бактериофагах и других объектах. В результате своих работ Бензер ввел три новых понятия:
- Ранее считали, что кроссинговер может происходить только между генами и, таким образом, ген - это элементарная единица генетической рекомбинации. Однако доказано, что рекомбинации происходят и внутри гена. Наименьшая единица рекомбинации названа реконом.
- Ранее считали ген единицей мутации. Однако было обнаружено, что изменения отдельных участков внутри сложного гена приводят к изменению его функции. Мельчайшая единица, способная к изменению, была названа мутоном.
- Ген считали единицей функции. Многочисленные исследования показали, что функция гена может изменяться в зависимости от того, расположены ли два мутантных аллеля сложного гена в одной хромосоме, а их нормальные аллели в гомологичной (цис-положение), или мутантные аллели расположены в двух гомологичных хромосомах (трансположение). Единицу функции предложено называть цистроном.
Параллельная работа биохимиков и генетиков показала, что наименьшая величина рекона и мутона близка к величине одного или нескольких нуклеотидов. Цистрон же гомологичен участку ДНК, "кодирующему" синтез определенного полипептида, и содержит тысячу и более нуклеотидов.
Функционально-генетическая классификация генов
Существует несколько классификаций генов (аллельные и неаллельные, летальные и полулетальные гены и др.). Характеристики гена как единицы функции наследственного материала и системный принцип организации генотипа отражены в функционально-генетической классификации наследственных задатков
Структурными называются гены, контролирующие развитие конкретных признаков. Продуктом первичной активности гена является либо иРНК и далее полипептид, либо рРНК и тРНК. Таким образом, структурные гены содержат информацию об аминокислотных или нуклеотидных последовательностях макромолекул. Структурные гены трех подгрупп, приведенных в классификации, отличаются степенью плейотропного действия, причем выраженная плейотропия отличает гены второй и третьей подгрупп, которые активно функционируют во всех клетках. При их мутациях наблюдаются разнообразные и обширные нарушения развития организма. Неслучайно поэтому эти гены представлены в генотипе в количестве нескольких десятков копий и образованы среднеповторяющимися последовательностями ДНК.
Гены-модуляторы смещают в ту или иную сторону процесс развития признака или другие генетические явления, например частоту мутирования структурных генов. Часть структурных генов выполняет одновременно и роль модуляторов (см. пример "эффекта положения"). Другие гены-модуляторы, по-видимому, лишены каких-либо иных генетических функций. Появление таких генов в эволюции имело большое значение. Благодаря плейотропному действию многие структурные гены, наряду с благоприятным и необходимым для нормального развития организма действием, оказывают и нежелательные эффекты, которые снижают жизнеспособность особи. Неблагоприятное действие их ослабляется генами-модуляторами.
К регуляторным относятся гены, координирующие активность структурных генов, контролирующие время включения различных локусов в процессе индивидуального развития в зависимости от типа клеток многоклеточного организма, а также от состояния среды.
Молекулярно-биологические представления о строении и функционировании генов
Идеи молекулярной биологии к настоящему времени проникли во все отрасли науки о жизни и определили главные тенденции развития теоретической, экспериментальной и прикладной биологии. Молекулярная биология складывалась в ходе исследований физико-химических свойств и биологической роли нуклеиновых кислот и белков. Основы ее были заложены работами по генетике вирусов и фагов, химической природе наследственного материала, механизму биосинтеза белка, биологическому коду, закономерностям ультраструктурной организации клетки. В связи с этим молекулярную биологию можно определить как область изучения закономерностей структуры и изменений информационных макромолекул и участия их в фундаментальных процессах жизнедеятельности.
В области генетики молекулярная биология вскрыла химическую природу вещества наследственности, показала физикохимические предпосылки хранения в клетке информации и точного копирования ее для передачи в ряде поколений. ДНК большинства биологических объектов (от млекопитающих до бактериофага) содержит равные количества нуклеотидов с пуриновыми (аденин, гуанин) и пиримидиновыми (тимин, цитозин) азотистыми основаниями. Это означает, что объединение молекул ДНК в двойную спираль осуществляется закономерно, в соответствии с принципом комплементарности - адениловый нуклеотид связывается с тимидиловым нуклеотидом, а гуаниловый с цитидиловым (рис. 53). Такая конструкция делает возможным полуконсервативный способ редупликации ДНК. Вместе с тем вдоль биспирали ДНК пары А - Т и Г - Ц располагаются случайным образом - А + Т ≠ Г + Ц. Следовательно, путем независимого комбинирования нуклеотидов, различающихся по азотистому основанию, по длине молекул ДНК удается записать разнообразную информацию, объем которой пропорционален количеству нуклеиновой кислоты в клетке.
Согласно молекулярно-биологическим представлениям ген как единица функционирования наследственного материала характеризуется сложным строением. Многие детали тонкой структуры гена остаются неизвестными. Вместе с тем успехи современной науки в этой области достаточно велики, чтобы можно было нарисовать принципиальную модель функционирующего гена.



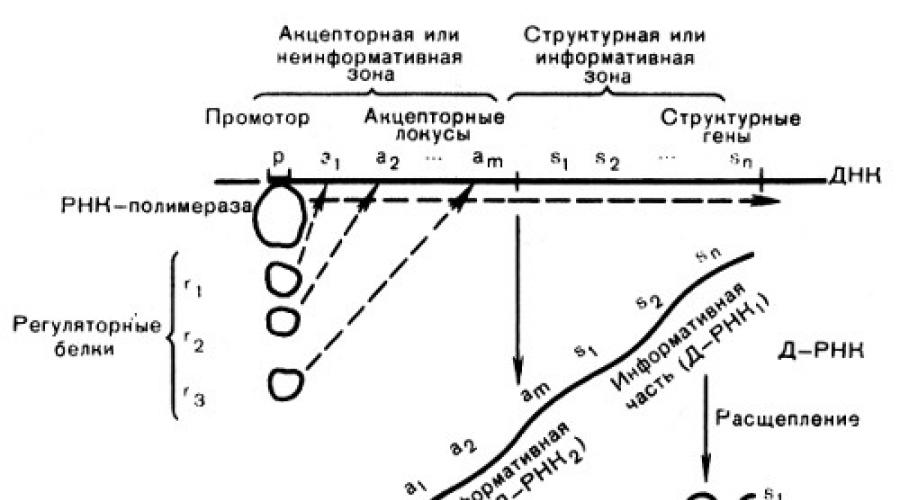
Функциональная активность гена заключается в синтезе на молекуле ДНК молекул РНК или транскрипции (переписывании) биологической информации с целью ее использования для образования белка. Единицы транскрипции (транскриптоны) превышают по размерам структурные гены (рис. 54). Согласно одной из моделей транскиптона в клетках эукариот он состоит из неинформативной (акцепторной) и информативной зоны. Последняя образована структурными генами (цистронами), которые разделены вставками ДНК - спейсерами, не несущими информации об аминокислотных последовательностях белков. Неинформативная зона начинается геном-промотором (р), к которому присоединяется фермент РНК-полимераза, катализирующая реакцию ДНК-зависимого образования рибонуклеиновых кислот. Далее следуют акцепторные гены или гены-операторы (α 1 , α 2 и т. д.), связывающие регуляторные белки (r 1 , r 2 ит. д.), изменения которых "открывают" ДНК структурных генов (s 1 , s 2 и т. д.) для считывания информации. На транскриптоне синтезируется одна большая молекула РНК. Благодаря процессингу неинформативная ее часть разрушается, а информативная расщепляется на фрагменты, соответствующие отдельным структурным генам. Эти фрагменты в виде иРНК для синтеза конкретных полипептидов транспортируются в цитоплазму. Согласно приведенной модели в транскрипте находится несколько структурных генов. Группа этих генов образует функциональный блок и называется опероном. Функциональное единство оперонов зависит от наличия генов-операторов, которые воспринимают сигналы из метаболического аппарата цитоплазмы и активируют структурные гены.
Природа сигналов, регулирующих функцию генов, изучена у прокариот. Это белки, синтез которых контролируется особыми генами-регуляторами, действующими на гены-операторы. Активация структурных генов посредством генов-регуляторов и операторов представлена на схеме (рис. 55). В обычных условиях ген-регулятор активен и в клетке протекает синтез белка-репрессора, который связывается с геном-оператором и блокирует его. Это выключает из функции весь оперон.
Включение оперона происходит, если в цитоплазму проникают молекулы субстрата, для переваривания которого требуется возобновление синтеза соответствующего фермента. Субстрат присоединяется к репрессору и лишает его способности блокировать ген-оператор. В этом случае информация со структурного гена считывается и требуемый фермент образуется. В описанном примере субстрат играет роль индуктора (побудителя) синтеза "своего" фермента. Последний запускает биохимическую реакцию, в которой используется данный субстрат. По мере снижения его концентрации освобождаются молекулы репрессора, которые блокируют активность гена-оператора, что приводит к выключению оперона. У бактерий описана система регуляции, переводящая активные структурные гены в неактивное состояние в зависимости от концентрации в цитоплазме конечного продукта определенной биохимической реакции (рис. 56). При этом под генетическим контролем гена-регулятора образуется неактивная форма репрессора гена-оператора. Репрессор активируется в результате взаимодействия с конечным продуктом данной биохимической реакции и, блокируя ген-оператор, выключает соответствующий оперон. Синтез фермента, катализирующего образование вещества, активирующего репрессор, прекращается. Описанные системы регуляции функции структурных генов носят приспособительный характер. В первом примере синтез фермента запускается поступлением в клетку субстрата соответствующей реакции, во втором - образование фермента прекращается, как только исчезает потребность в синтезе определенного вещества.
Принципы регуляции генной активности у эукариот, по-видимому, сходны с таковыми у бактерий. Вместе с тем появление ядерной оболочки, усложнение генных взаимодействий в условиях диплоидности, необходимость тонкой корреляции генетических функций отдельных клеток многоклеточного организма повлекли за собой при переходе к эукариотическому типу клеточной организации усложнение регуляторно-генетических механизмов, генетико-биохимические и кибернетические основы которых во многом еще не выяснены. Можно предположить также, что в эволюции увеличилось число генов-операторов. Индукторами транскрипции многих структурных генов эукариот служат гормоны. Предполагается наличие генов-интеграторов, включающих в ответ на стимул одновременно "батареи генов". Генетическая система высших организмов отличается, по-видимому, большой гибкостью реакций на действие негенетических факторов. В подтверждение этого допущения рассмотрим ряд факторов. Так, некоторые структурные гены животных не являются непрерывными последовательностями кодонов, а составлены из фрагментов, которые прерываются неинформативными участками ДНК. Ген Р-полипептида гемоглобина мыши, например, прерывается вставкой из 550 пар нуклеотидов. Соответствующий этой вставке участок отсутствует в зрелой глобиновой иРНК, что говорит о его разрушении в ходе процессинга первичной транскрибированной РНК с воссоединением информационных фрагментов иРНК. Информационные участки таких генов получили название экзонов, "молчащие" - интронов, а процесс воссоединения информационных фрагментов иРНК - сплайсинга (сплавления). Количество ДНК в области нитронов в 5-10 раз выше, чем в области экзонов. Предполагается, что сплайсинг служит механизмом образования некоторых генов в момент их функциональной активности, т. е. на 1 уровне иРНК.
Известны также "блуждающие" структурные гены, положение которых в хромосоме меняется в зависимости от фазы жизненного цикла. Так, "тяжелые" и "легкие" полипептиды иммуноглобулинов состоят из константного (С) и вариабельного (Y) участков, синтез которых контролируется сцепленными, но разными генами. В зрелых плазматических клетках эти гены разделены нетранскрибируемой вставкой длиной в 1000 пар нуклеотидов. В клетках эмбрионов названная вставка во много раз длиннее. Таким образом в процессе клеточной дифференцировки изменяется взаиморасположение генов. Исследование механизмов регуляции генной активности и генных взаимодействий у эукариот представляет важнейшую область современной молекулярной биологии и генетики.
Свойства гена
Ген как единица функционирования наследственного материала имеет ряд свойств.
- Специфичность - уникальная последовательность нуклеотидов для каждого структурного гена, т.е. каждый ген кодирует свой признак;
- Целостность - как функциональная единица (программирование синтеза белка) ген неделим;
- Дискретность - в составе гена имеются субъединицы: мутон - субъединица, отвечающая за мутацию, рекон - отвечает за рекомбинаци. Минимальная их величина - пара нуклеотидов;
- Стабильность - ген, как дискретная единица наследственности отличается стабильностью (постоянством) - при отсутствии мутации он передается в ряду поколений в неизменном виде. Частота самопроизвольной мутации одного гена составляет примерно 1·10 -5 на поколение.
- Лабильность - устойчивость генов не абсолютная, они могут изменяться, мутировать;
- Плейотропия - множественный эффект отдельного гена (один ген отвечает за несколько признаков);
Примером плейотропного эффекта гена у человека служит синдром Марфана. Хотя это наследственное заболевание зависит от присутствия в генотипе одного измененного гена, оно характеризуется в типичных случаях триадой признаков: подвывихом хрусталика глаза, аневризмой аорты, изменениями опорно-двигательного аппарата в виде "паучьих пальцев", деформированной грудной клеткой, высоким сводом стопы. Все перечисленные признаки являются сложными. По-видимому, в основе их лежит один и тот же дефект развития соединительной ткани.
Так как продуктом функции гена наиболее часто является белок-фермент, выраженность плейотропного эффекта зависит от распространенности в организме биохимической реакции, которую катализирует фермент, синтезируемый под генетическим контролем данного гена. Распространенность поражений в организме в случае наследственного заболевания тем больше, чем выраженнее плейотропный эффект измененного гена.
Ген, имеющийся в генотипе в необходимом для проявления количестве (1 аллель для доминантных признаков и 2 аллеля для рецессивных), может проявляться в виде признака в разной степени у разных организмов (экспрессивность) или вообще не проявляться (пенетрантность). Экспрессивность и пенетрантность определяются факторами среды (воздействием условий окружающей среды - модификационной изменчивостью) и влиянием других генов генотипа (комбинативная изменчивость).
- Экспрессивность - степень выраженности гена в признаке или степень фенотипического проявления гена.
Например, аллели групп крови АВ0 у человека имеют постоянную экспрессивность (всегда проявляются на 100%), а аллели, определяющие окраску глаз, – изменчивую экспрессивность. Рецессивная мутация, уменьшающая число фасеток глаза у дрозофилы, у разных особей по разному уменьшает число фасеток вплоть до полного их отсутствия.
- Пенетрантность - частота фенотипического проявления признака при наличии соответствующего гена (отношение (в процентах) числа особей, имеющих данный
признак, к числу особей, имеющих данный ген);
Например, пенетрантность врожденного вывиха бедра у человека составляет 25%, т.е. болезнью страдает только 1/4 рецессивных гомозигот. Медико-генетическое значение пенетрантности: здоровый человек, у которого один из родителей страдает заболеванием с неполной пенетрантностью, может иметь непроявляющийся мутантный ген и передать его детям.
– РНК), определяющий (кодирующий) возможность развития какого-либо признака. Ген – функционально неделимая единица, т. е. один ген, как правило, отвечает за один элементарный признак. Таким признаком на молекулярном уровне может быть молекула белка или РНК, а на уровне организма, напр., цвет или цвет глаз человека. Вместе с тем возможность реализации гена, его проявления в виде признака зависят от ряда факторов, прежде всего от взаимодействия с другими генами, образующими среду (см. Генотип).
Изучение строения, организации, принципов работы генов (или несколько шире – генетического материала) – центральная проблема генетики на всех этапах её развития. При этом представления о гене как о наследственном факторе, обладающем функцией, физической природой, способностью к изменчивости и другими свойствами, существенно изменялись и дополнялись. В 1865 г. Г. Мендель на основании своих опытов по растений доказал существование дискретных наследственных «задатков», которые датский генетик В. Иогансен в 1909 г. назвал генами. Работы Менделя открыли возможность точного генетического () анализа наследственности и после их повторения в 1900 г. дали толчок необычайно быстрому становлению генетики. Уже в первой трети 20 в. было установлено, что гены линейно расположены в хромосомах клеточного ядра (см. Хромосомная теория наследственности), что они могут подвергаться естественным или вызываемым искусственно наследуемым изменениям – мутациям и что при передаче их от родителей к потомкам происходит их перераспределение – рекомбинация. При этом оказалось, что ген как единица функции и ген как единица мутации и рекомбинации – не одно и то же. Так возникло представление о сложном строении гена, однако вопрос о его химической природе оставался нерешённым. Наконец, в 40-х гг. на микроорганизмах было показано, что веществом генов является дезоксирибонуклеиновая кислота (ДНК), а в 1953 г. создана её пространственная модель (т. н. двойная спираль), объяснявшая биологические функции этой гигантской молекулы её строением. Началось бурное развитие молекулярной биологии гена. Вскоре были раскрыты способы записи генетиче-ской информации (генетический код) и механизм её передачи в процессах репликации, транскрипции и трансляции. Ещё в 40-х гг. была выдвинута концепция: «один ген – один фермент», согласно которой каждый ген определяет структуру какого-либо фермента (белка). Теперь это положение уточнялось: если белок состоит из нескольких полипептидных цепей, то каждая из них кодируется отдельным геном, т. е. более верна формула: «один ген – одна полипептидная цепь». В клетках существуют набор генов, специфичный для организмов одного биологического вида, и механизмы регуляции их активности. Благодаря этому происходит регулируемый синтез ферментов и других белков, обеспечивающих специализацию клеток и тканей в процессе развития организма из оплодотворённой яйцеклетки и поддерживающих характерный для вида тип обмена веществ.
В дальнейшем были исследованы особенности организации генетического материала у прокариот, эукариот и вирусов, а также у клеточных органоидов – митохондрий и хлоропластов, открыты т. н. мобильные гены, перемещающиеся по , расшифрована структура (нуклеотидная последовательность) геномов ряда организмов, в т. ч. человека. Разработка методов выделения, клонирования и гибридизации отдельных генов (участков ДНК) привела к появлению важной в практическом отношении генной инженерии, ряда направлений в биотехнологии. См. также Аллель, Геном, Хроматин.
Дискретной единицей наследственности у высших организмов является ген. Совокупность всех генов определенного биологического вида определяется термином геном (иногда данный термин относится к полной генетической системе отдельной клетки или конкретного организма). Ген в своем наиболее практическом понимании представляет собой строго определенный участок молекулы ДНК, последовательность которого заключает в себе всю информацию, необходимую для синтеза молекулы белка или РНК. Генетическая информация зашифрована посредством универсального для всех живых организмов генетического кода, представляющего собой набор нуклеотидных триплетов - кодонов. Каждый такой триплет (т.е. каждая последовательность из 3 нуклеотидов) кодирует синтез одной, строго определенной аминокислоты в составе белка.
Считывание кодонов в процессе передачи генетической информации происходит последовательно (принцип линейности генетического кода), и любой нуклеотид может входить в состав только одного кодона (принцип неперекрываемости генетического кода). Генетический код является вырожденным, т.е. допускает кодирование каждой из 20 аминокислот несколькими г.озможными комбинациями триплетов (всего таких комбинаций может быть 64). Расшифровка точной последовательности нуклеотидов определенного информационного участка гена позволяет однозначно идентифицировать последовательность аминокислот в составе соответствующего полипептидного участка белка и его размер. Полный гаплоидный геном человека (т.е. кодируемый одной смысловой нитью ДНК) включает, ориентировочно, около 30 000-40 000 генов.
Гены человека и других высших организмов
имеют чрезвычайно сложную структурно-функциональную организацию и содержат различные по своей биологической роли нуклеотидные участки. Одни из них (экзоны) являются относительно короткими, представляют собой кодирующие последовательности и определяют аминокислотный состав белков; другие участки гена (интроны) являются обычно значительно более протяженными и не несут непосредственной информационной нагрузки. Окончательная роль интронов до настоящего времени не установлена; предполагается, что они могут иметь отношение к регуляции экспрессии генов и контролю тонких механизмов «считывания» генетической информации. В состав генов входят также особые регу-ляторные участки (промоторы, энхансеры, различные сигнальные последовательности), обеспечивающие инициацию, интенсивность и определенную временную последовательность процессов нуклеотидного синтеза на ДНК-матрице, а также модификацию промежуточных полинуклеотидных продуктов.
По ориентировочным оценкам
, собственно кодирующие последовательности ДНК составляют не более 3-10% всего генома человека.
В любой клетке организма содержится полный набор генов, однако лишь небольшая их часть является функционально активной в каждой конкретной ткани, т.е. экспрессируется. Под экспрессией гена понимают реализацию записанной в нем генетической информации, приводящую к синтезу первичных молекулярных продуктов гена - РНК и белка. Именно временная и тканевая избирательность экспрессии генов определяет специфику дифференцировки и функционирования различных органов, тканей и клеток организма в онтогенезе.
Ген
I
(греч. genos , происхождение)
структурно-функциональная единица генетического материала, наследственный фактор, который можно условно представить как отрезок молекулы (у некоторых вирусов - молекулы ), включающий нуклеотидную последовательность, в которой закодирована первичная структура полипептида (белка) либо молекулы транспортной или рибосомной РНК, которых контролируется этим . Обусловливая первичную структуру конкретного белка, ген тем самым определяет формирование отдельного признака организма или клетки. Предположение о существовании наследственных факторов впервые было высказано Менделем (G.J. Mendel) в 1865 г., который пришел к заключению, что передача признака родителей потомству обусловлена передачей через этих наследственных факторов, каждый из которых передается как нечто целое и независимое. В 1909 г. Иоганнсен (W. Johannsen) предложил обозначать менделевские наследственные факторы термином «гены». В 1911 г. Морганом (Th.Н. Morgan) и его сотрудниками было показано, что ген является участком и что отдельная состоит из генов, последовательно расположенных по ее длине (см. Хромосомы).
Каждый ген занимает свое определенное место () на хромосоме.
Позднее Морганом и его сотрудниками были созданы первые хромосомные карты, на которых они показали расположение отдельных генов на хромосомах. Совокупность хромосомных (или ядерных) генов, составляющих так называемый геном, и генов, локализованных в цитоплазматических структурах - митохондриях, пластидах, плазмидах, определяет клетки или организма. Ген может непосредственно определять наличие какого-либо признака (фена) организма или принимать участие в формировании нескольких признаков (явление плейотропии). Однако основная масса признаков у человека формируется в результате взаимодействия многих генов (явление полигении). Утрата гена или его изменение (см. Мутагенез) приводят к изменению признака, контролируемого этим геном. Степень проявления признака, контролируемого конкретным геном ( гена), зависит также от условий окружающей среды. В то же время даже в пределах родственной группы особей, находящихся в сходных условиях существования, проявление одного и того же гена может варьировать по степени выраженности. Все это свидетельствует о том, что при формировании признаков генотип выступает как целостная , функционирующая в строгой зависимости от внутриорганизменной и окружающей среды. Т.о., отдельный признак или совокупность всех признаков организма, т.е. его , являются результатом взаимодействия генотипа с окружающей средой; способность гена фенотипически проявлять себя тем или иным образом называют пенетрантностью гена. У диплоидных организмов, т.е. у организмов, соматические клетки которых имеют , гены представлены парой аллелей. Аллель - это одно из возможных состояний или один из возможных вариантов гена; теоретически число аллелей каждого гена неисчислимо, но не все они прошли эволюционный . В гомологичных хромосомах аллельные гены расположены в гомологичных локусах. Аллельная генов может быть составлена из идентичных (явление гомозиготности) или различных (явление гетерозиготности) аллелей. У гетерозигот (организмов, аллельные гены которых различны) проявление одного аллеля на уровне признака организма (фенотипическое проявление) может полностью подавлять проявление другого аллеля. Подавляющий аллель называют доминантным, а подавляемый - рецессивным. Соответственно и контролируемые ими признаки носят название доминантных или рецессивных. Фенотипическое проявление рецессивных генов можно наблюдать только у тех организмов, которые оказываются гомозиготными в отношении такого рецессивного гена, т.е. оба аллельных гена у них рецессивны, или в случае, когда ген не имеет аллельной пары, например некоторые гены, расположенные на одной из половых хромосом при их XY-сочетании. У гетерозиготных организмов возможно и совместное (кодоминантное) проявление аллелей. Т.о., понятия « » и «рецессивный» отражают вклад данного гена в формирование конкретного признака. Свойство гена подавлять или быть подавленным в значительной мере зависит также от генного окружения - генотипической среды, в которой находится этот ген. Перенос гена в другое место хромосомы, влекущий за собой изменение его генного окружения, ведет к утрате этим геном своих свойств, в т.ч. даже такого свойства, выработанного в процессе длительной эволюции, как способность доминировать. Это явление называют эффектом положения гена. При возвращении гена в прежнее положение на хромосоме его способность доминировать восстанавливается. Изучая механизмы регуляции функции гена, французские генетики Жакоб (F. Jacob) и (J.L. Monod) пришли к заключению, что существуют структурные и регуляторные гены. К структурным генам относятся гены, которые контролируют (кодируют) первичную структуру матричных, или информационных, РНК, а через них последовательность аминокислот в синтезируемых полипептидах (см. Белки).
Другую группу структурных генов составляют гены, определяющие последовательность нуклеотидов в полинуклеотидных цепях рибосомной РНК и транспортной РНК (см. Нуклеиновые кислоты).
Регуляторные гены контролируют синтез специфических веществ, так называемых ДНК-связывающих белков, которые регулируют структурных генов. Используя способность некоторых Бактериофаг ов переносить фрагменты бактериальной хромосомы в другие бактериальные клетки (явление трансдукции), Беквит (J.R. Beckwith) и его сотрудники в 1969 г. впервые выделили, точно определили размер индивидуального гена кишечной палочки и получили его электронограмму. В 1967-1970 гг. Корана (Н.G. Khorana) осуществил химический синтез индивидуального гена. По мере увеличения возможностей генетического анализа (см. Генетика) были получены все новые доказательства того, что ген, являясь функциональной единицей, вместе с тем имеет весьма сложное строение. Первые доказательства сложности организации гена получили в 1929 г. советские ученые А.С. Серебровский, Н.П. Дубинин и И.И. Агол. Наряду со структурными и регуляторными генами в молекулах ДНК были обнаружены участки повторяющихся нуклеотидных последовательностей, функции которых не известны, а также мигрирующие нуклеотидные последовательности - так называемые мобильные гены. Найдены также псевдогены, представляющие собой неактивные копии известных генов, но расположенные в других частях генома. В 1953 г.
английский биохимик Крик (F. Н.С. Crick) и американский биохимик Уотсон (J.D. Watson) предложили строения молекулы ДНК и высказали предположение, вскоре полностью подтвердившееся, что последовательность нуклеотидов в полинуклеотидной цепи ДНК является кодом, в соответствии с которым осуществляется соединение аминокислотных остатков в полипептидной цепи белковых молекул, строящихся под контролем соответствующих генов. В дальнейшем этот генетический был изучен более подробно. Было установлено, что включение одного аминокислотного остатка в строящуюся полипептидную цепь определяется сочетанием трех последовательно расположенных нуклеотидов, так называемых триплетов, причем включение одной и той же могут кодировать несколько различных триплетов Доказано, что генетический код универсален, т.е. он един для всех живых организмов. Реализация информации, «записанной» в гене, осуществляется с помощью посредника, которым является одна из разновидностей РНК - матричная, или информационная, РНК (). мРНК происходит на молекуле ДНК как на матрице. Такой матричный синтез обеспечивает точность «переписывания» (транскрипции) особенностей нуклеотидной последовательности гена на молекулу мРНК. Синтезированная мРНК из ядра клетки поступает в цитоплазму, где на рибосомах (см. Клетка) происходит реализация генетической информации (процесс трансляции), которая воплощается в последовательность аминокислот, соединяющихся в полипептидную цепь белка. Средняя по размерам молекула белка содержит около 300 аминокислотных остатков. Следовательно, средний ген должен содержать не менее 1000-1500 нуклеотидов. Однако количество нуклеотидных в обычной молекуле ДНК по крайней мере в 10 раз превышает количество генов. Такая «избыточность» ДНК объясняется тем, что, например, у человека только 6-10% всей ДНК составляют кодирующие специфические нуклеотидные последовательности, остальные нуклеотиды в генетическом кодировании непосредственно не участвуют. Большинство генов эукариот имеет прерывистую структуру: участок ДНК, кодирующий аминокислотную последовательность полипептидной цепи белка, разделен некодирующими вставками на несколько частей. Кроме того, некоторые некодирующие нуклеотидные последовательности обрамляют транскрибируемую единицу с концов. При транскрипции и те, и другие участки ДНК «считываются» в виде единой молекулы-предшественницы мРНК. Затем некодирующие участки выщепляются, а кодирующие участки соединяются друг с другом, образуя молекулу «зрелой» мРНК, способной транслироваться в молекулу белка. Другие некодирующие нуклеотидные последовательности могут играть роль сигнальных последовательностей, ответственных за начало определенных процессов в клетке. К ним относятся так называемые промоторы транскрипции, точки начала репликации ДНК, участки скручивания хромосом и др. Некодирующие последовательности состоят из множества семейств, характеризующихся разной степенью повторяемости нуклеотидов и различной организацией. Однако только немногие из этих последовательностей изучены настолько, чтобы определенной последовательности могла быть приписана определенная . Т.о., ген представляет собой сложную микросистему, обеспечивающую жизнедеятельность клетки и организма в . Теория гена, постоянно углубляющаяся и развивающаяся, является основой генетической инженерии (Генетическая инженерия),
конечной целью которой служит создание организмов с новыми наследственными свойствами, а также разработка способов лечения генетически обусловленных заболеваний (см. Наследственные болезни).
структурная и функциональная единица наследственности, контролирующая образование какого-либо признака, представляющая собой отрезок молекулы дезоксирибонуклеиновой кислоты (у некоторых вирусов - рибонуклеиновой кислоты). Ген амбивале́нтный
(лат. приставка ambi- вокруг, с обеих сторон + valens, valentis сильный) - Г., оказывающий как полезное, так и вредное действие на его носителя. Ген аутосо́мный
- Г., локализованный в любой хромосоме, за исключением половых. Ген внехромосо́мный
( . Г. нехромосомный) - Г., локализованный вне хромосом в той или иной цитоплазматической структуре. Ген голандри́ческий
(греч. holos весь, полностью + anēr, andros мужчина) - Г., локализованный в участке Y-хромосомы, не имеющем гомологии в X-хромосоме, и поэтому абсолютно сцепленный с Y-хромосомой. Ген гомеоти́ческий
(греч. homoios подобный) - Г., действие которого обусловливает трансформацию эмбрионального зачатка одного органа в другой, возникающий обычно в несвойственном ему месте. Ге́ны гомодинами́ческие
- Г., контролирующие одновременно одни и те же процессы развития. Ге́ны гомологи́чные
- Г. особей одного и того же биологического вида или разных видов с одинаковой функцией и локализацией относительно других генов. Ген диагини́ческий
(греч. dia через + gynē женщина) - Г. X-хромосомы, переданной от матери к сыну. Ген диандри́ческий
(греч. dia через + anēr, andros мужчина) - Г. X-хромосомы, переданной от отца к дочери. Ген домина́нтный
(лат. dominans, dominantis господствующий) - Г., сходно проявляющийся в гетеро- и гомозиготном состоянии и подавляющий проявление других аллелей этого гена. Ген зави́симый
(син. Г. криптомерный - устар.) - Г., контролирующий при полигении образование специфического признака лишь во взаимодействии с другими неаллельными генами. Ген идиомо́рфный
(греч. idios своеобразный, необычный + morphē , форма) - Г., у которого один аллель заполняет всю популяцию, а все другие вместе встречаются с частотой, не превышающей 1%. Ген изоляцио́нный
- Г., в гетерозиготном состоянии обусловливающий снижение жизнеспособности или плодовитости особи. Ге́ны комбинацио́нные
- Г., детерминирующие различные процессы развития особи и образующие вторичный признак только путем комбинированного действия. Ге́ны компенсацио́нные
- как правило, рецессивные Г., взаимно изменяющие фенотипическое проявление друг друга. Ген ко́мплексный
- Г., состоящий из частей, контролирующих один и тот же признак, не могущих быть разделенными при кроссинговере. Ге́ны комплемента́рные
(лат. complementum дополнение) - неаллельные Г., каждый из которых может по-разному изменять один и тот же признак. Ген, контроли́руемый по́лом
(син. Г., модифицированный полом) - Г., присутствующий в генотипе обоих полов, но проявляющийся по-разному у особей мужского и женского пола. Ген криптоме́рный
(устар.; греч. kryptos скрытый + meros часть) - см. Ген зависимый. Ген лаби́льный
- Г., переходящий из одного стабильного состояния в другое через ряд мелких мутационных изменений. Ген лаби́льный в разви́тии
- Г., проявление которого сильно варьирует или отмечается не у всех особей. Ген лаби́льный к среде́
- Г., проявление которого в значительной степени зависит от условий окружающей и внутренней среды. Ген лета́льный
- Г., обусловливающий гибель особи обычно до достижения ею половой зрелости. Ген «межвидово́й
» - Г., детерминирующий межвидовые барьеры и не передающийся при межвидовом скрещивании. Ге́ны мно́жественные
- см. Гены полимерные. Ген, модифици́рованный по́лом
- см. Ген, контролируемый полом. Ген мута́бельный
(лат. mutabilis изменчивый) - Г., отличающийся высокой частотой спонтанного мутирования. Ге́ны неалле́льные
- Г., занимающие неидентичные локусы хромосом. Ген незави́симый
- Г., в случае полигении способный самостоятельно детерминировать образование признака без участия других генов, контролирующих этот признак. Ген нехромосо́мный
- см. Ген внехромосомный. Ген, ограни́ченный по́лом
- Г., присутствующий у особей обоих полов, но фенотипически проявляющийся только у особей одного пола. Ген плазмочувстви́тельный
- локализованный в хромосоме Г., проявление которого зависит от действия внехромосомных Г. Ген плейотро́пный
(греч. pleiōn более многочисленный + tropos направление) - Г., принимающий участие в формировании одновременно нескольких признаков. Ге́ны полиме́рные
(греч. polymerēs состоящий из многих частей, множественный; син.: , Г. множественные, ) - неаллельные Г., принимающие участие в формировании одного и того же признака. Ге́ны полиплика́тные
(греч. poly- много + лат. plico, plicatum складывать) - идентичные пары Г. с одинаковым фенотипическим проявлением, но локализованные в разных хромосомах; различают дупликатные, трипликатные, квадрипликатные Г. и т.д., соответственно числу таких пар. Ген полиурги́ческий
(греч. poly- много + греч. ergon действие) - Г., вызывающий неодинаковый эффект в различных частях организма соответственно специфическим свойствам протоплазмы. Ген регулято́рный
- Г., осуществляющий контроль активности оперона. Ген рецесси́вный
- Г., проявляющийся только в гомозиготном состоянии. Ген сигна́льный
(син. ген-маркер) - Г. с известной локализацией и проявлением, используемый для картирования данной хромосомы. Ген сло́жный
- Г., состоящий из частей, не разделяемых кроссинговером, но обладающих независимой мутабельностью и частично независимых друг от друга. Ген, стаби́льный в разви́тии
- Г., характеризующийся регулярным и не варьирующим по силе проявлением. Ген, сце́пленный с по́лом
- Г., локализованный в половой хромосоме; различают Г., абсолютно и неполностью сцепленные с полом. Ге́ны цепны́е
- группа Г., каждый из которых контролирует прохождение отдельного этапа в цепи реакций, обусловливающих в итоге образование признака. Ге́ны эквилока́льные
(лат. aequus равный, одинаковый + locus место, положение) - Г., занимающие идентичные участки гомологичных хромосом. 1. Малая медицинская энциклопедия. - М.: Медицинская энциклопедия. 1991-96 гг. 2. Первая медицинская помощь. - М.: Большая Российская Энциклопедия. 1994 г. 3. Энциклопедический словарь медицинских терминов. - М.: Советская энциклопедия. - 1982-1984 гг
.
"хромосома" - слова, которые знакомы каждому школьнику. Но представление об этом вопросе довольно обобщенное, так как для углубления в биохимические дебри требуются специальные знания и желание все это понимать. А оно, если и присутствует на уровне любопытства, то быстро пропадает под тяжестью изложения материала. Попробуем разобраться в хитросплетениях в научно-полярной форме.
Ген - это наименьшая структурная и функциональная частица информации о наследственности у живых организмов. По сути он представляет собой небольшой участок ДНК, в котором содержится знание об определенной последовательности аминокислот для построения белка либо функциональной РНК (с которой также будет синтезирован белок). Ген определяет те признаки, которые будут наследоваться и передаваться потомками дальше по генеалогической цепи. У некоторых одноклеточных организмов существует перенос генов, который не имеет отношения к воспроизведению себе подобных, он называется горизонтальным.
"На плечах" генов лежит огромная ответственность за то, как будет выглядеть и работать каждая клетка и организм в целом. Они управляют нашей жизнью от момента зачатия до самого последнего вздоха.

Первый научный шаг вперед в изучении наследственности был сделан австрийским монахом Грегором Менделем, который в 1866 году опубликовал свои наблюдения о результатах при скрещивании гороха. Наследственный материал, который он использовал, четко показывал закономерности передачи признаков, таких как цвет и форма горошин, а также цветки. Этот монах сформулировал законы, которые сформировали начало генетики как науки. Наследование генов происходит потому, что родители отдают своему чаду по половинке всех своих хромосом. Таким образом, признаки мамы и папы, смешиваясь, образуют новую комбинацию уже имеющихся признаков. К счастью, вариантов больше, чем живых существ на планете, и невозможно отыскать двух абсолютно идентичных существ.
Мендель показал, что наследст-венные задатки не смешиваются, а передаются от родителей потомкам в виде дискретных (обособлен-ных) единиц. Эти единицы, представленные у особей парами (аллелями), остаются дискретными и передаются по-следующим поколениям в мужских и женских га-метах, каждая из которых содержит по одной едини-це из каждой пары. В 1909 году датский ботаник Иогансен назвал эти единицы генами. В 1912 году генетик из Соединенных Штатов Америки Морган показал, что они находятся в хромосомах.
С тех пор прошло больше полутора веков, и исследования продвинулись дальше, чем Мендель мог себе представить. На данный момент ученые остановились на мнении, что информация, находящаяся в генах, определяет рост, развитие и функции живых организмов. А может быть, даже и их смерть.
Классификация
Структура гена содержит в себе не только информацию о белке, но и указания, когда и как ее считывать, а также пустые участки, необходимые для разделения информации о разных белках и остановки синтеза информационной молекулы.
Существует две формы генов:
- Структурные - они содержат в себе информацию о строении белков или цепей РНК. Последовательность нуклеотидов соответствует расположению аминокислот.
- Функциональные гены отвечают за правильную структуру всех остальных участков ДНК, за синхронность и последовательность ее считывания.
На сегодняшний день ученые могут ответить на вопрос: сколько генов в хромосоме? Ответ вас удивит: около трех миллиардов пар. И это только в одной из двадцати трех. Геном называется наименьшая структурная единица, но она способна изменить жизнь человека.
Мутации

Случайное или целенаправленное изменение последовательности нуклеотидов, входящих в цепь ДНК, называется мутацией. Она может практически не влиять на структуру белка, а может полностью извратить его свойства. А значит, будут локальные или глобальные последствия такого изменения.
Сами по себе мутации могут быть патогенными, то есть проявляться в виде заболеваний, либо летальными, не позволяющими организму развиваться до жизнеспособного состояния. Но большинство изменений проходит незаметно для человека. Делеции и дупликации постоянно совершаются внутри ДНК, но не влияют на ход жизни каждого отдельного индивидуума.
Делеция - это потеря участка хромосомы, который содержит определенную информацию. Иногда такие изменения оказываются полезными для организма. Они помогают ему защититься от внешней агрессии, например вируса иммунодефицита человека и бактерии чумы.
Дупликация - это удвоение участка хромосомы, а значит, и совокупность генов, которые он содержит, также удваивается. Из-за повторения информации она хуже подвержена селекции, а значит, может быстрее накапливать мутации и изменять организм.
Свойства гена
У каждого человека имеется огромная Гены - это функциональные единицы в ее структуре. Но даже такие малые участки имеют свои уникальные свойства, позволяющие сохранять стабильность органической жизни:
- Дискретность - способность генов не смешиваться.
- Стабильность - сохранение структуры и свойств.
- Лабильность - возможность изменяться под действием обстоятельств, подстраиваться под враждебные условия.
- Множественный аллелизм - существование внутри ДНК генов, которые, кодируя один и тот же белок, имеют разную структуру.
- Аллельность - наличие двух форм одного гена.
- Специфичность - один признак = один ген, передающийся по наследству.
- Плейотропия - множественность эффектов одного гена.
- Экспрессивность - степень выраженности признака, который кодируется данным геном.
- Пенетрантность - частота встречаемости гена в генотипе.
- Амплификация - появление значительного количества копий гена в ДНК.
Геном

Геном человека - это весь наследственный материал, который находится в единственной клетке человека. Именно в нем содержатся указания о построении тела, работе органов, физиологических изменениях. Второе определение данного термина отражает структуру понятия, а не функцию. Геном человека - это совокупность генетического материала, упакованного в гаплоидном наборе хромосом (23 пары) и относящегося к конкретному виду.
Основу генома составляет молекула хорошо известная как ДНК. Все геномы содержат по крайней мере два вида информации: кодированная информация о структуре молекул-посредников (так называемых РНК) и белка (эта информация содержится в генах), а также инструкции, которые определяют время и место проявления этой информации при развитии организма. Сами гены занимают небольшую часть генома, но при этом являются его основой. Информация, записанная в генах, — своего рода инструкция для изготовления белков, главных строительных кирпичиков нашего тела.
Однако для полной характеристики генома недостаточно заложенной в нем информации о структуре белков. Нужны еще данные об элементах которые принимают участие в работе генов, регулируют их проявление на разных этапах развития и в разных жизненных ситуациях.
Но даже и этого мало для полного определения генома. Ведь в нем присутствуют также элементы, способствующие его самовоспроизведению (репликации), компактной упаковке ДНК в ядре и еще какие-то непонятные пока еще участки, иногда называемые «эгоистичными» (то есть будто бы служащими только для самих себя). По всем этим причинам в настоящий момент, когда заходит речь о геноме, обычно имеют в виду всю совокупность последовательностей ДНК, представленных в хромосомах ядер клеток определенного вида организмов, включая, конечно, и гены.
Размер и структура генома

Логично предположить, что ген, геном, хромосома отличаются у разных представителей жизни на Земле. Они могут быть как бесконечно маленькими, так и огромными и вмещать в себе миллиарды пар генов. Структура гена также будет зависеть от того, чей геном вы исследуете.
По соотношению между размерами генома и числом входящих в него генов можно выделить два класса:
- Компактные геномы, имеющие не более десяти миллионов оснований. У них совокупность генов строго коррелирует с размером. Наиболее характерны для вирусов и прокариотов.
- Обширные геномы состоят более чем из 100 миллионов пар оснований, не имеющих взаимосвязи между их длиной и количеством генов. Чаще встречаются у эукариотов. Большинство нуклеотидных последовательностей в этом классе не кодируют белков или РНК.
Исследования показали, что в геноме человека находится около 28 тысяч генов. Они неравномерно распределены по хромосомам, но значение этого признака остается пока загадкой для ученых.
Хромосомы

Хромосомы - это способ упаковки генетического материала. Они находятся в ядре каждой эукариотической клетки и состоят из одной очень длинной молекулы ДНК. Их легко можно увидеть в световой микроскоп в процессе деления. Кариотипом называется полный набор хромосом, который является специфичным для каждого отдельного вида. Обязательными элементами для них являются центромера, теломеры и точки репликации.
Изменения хромосом в процессе деления клетки
Хромосома - это последовательные звенья цепи передачи информации, где каждое следующее включает предыдущее. Но и они претерпевают определенные изменения в процессе жизни клетки. Так, например, в интерфазе (период между делениями) хромосомы в ядре расположены рыхло, занимают много места.
Когда клетка готовится к митозу (т. е. к процессу разделения надвое), хроматин уплотняется и скручивается в хромосомы, и теперь его становится видно в световой микроскоп. В метафазе хромосомы напоминают палочки, близко расположенные друг к другу и соединенные первичной перетяжкой, или центромерой. Именно она отвечает за формирование веретена деления, когда группы хромосом выстраиваются в линию. В зависимости от размещения центромеры существует такая классификация хромосом:
- Акроцентрические - в этом случае центромера расположена полярно по отношению к центру хромосомы.
- Субметацентрические, когда плечи (то есть участки, находящиеся до и после центромеры) неравной длины.
- Метацентрические, если центромера разделяет хромосому ровно посередине.
Данная классификация хромосом была предложена в 1912 году и используется биологами вплоть до сегодняшнего дня.
Аномалии хромосом

Как и с другими морфологическими элементами живого организма, с хромосомами тоже могут происходить структурные изменения, которые влияют на их функции:
- Анеуплоидия. Это изменение общего числа хромосом в кариотипе за счет добавления или удаления одной из них. Последствия такой мутации могут быть летальными для еще не родившегося плода, а также приводить к врожденным дефектам.
- Полиплоидия. Проявляется в виде увеличения количества хромосом, кратного половине их числа. Чаще всего встречается у растений, например водорослей, и грибов.
- Хромосомные аберрации, или перестройки, - это изменения в строении хромосом под воздействием факторов внешней среды.
Генетика
Генетика - это наука, изучающая закономерности наследственности и изменчивости, а также обеспечивающие их биологические механизмы. В отличие от многих других биологических наук она с момента своего возникновения стремилась быть точной наукой. Вся история генетики — это история создания и использования все более и более точных методов и подходов. Идеи и методы генетики играют важную роль в медицине, сельском хозяйстве, генетической инженерии, микробиологической промышленности.
Наследственность - способность организма обеспечивать в ряду морфологических, биохимических и физиологических признаков и особенностей. В процессе наследования воспроизводятся основные видоспецифические, групповые (этнические, популяционные) и семейные черты строения и функционирования организмов, их онтогенеза (индивидуального развития). Наследуются не только определенные структурно-функциональные характеристики организма (черты лица, некоторые особенности обменных процессов, темперамента и др.), но и физико-химические особенности строения и функционирования основных биополимеров клетки. Изменчивость — разнообразие признаков среди представителей определенного вида, а также свойство потомков приобретать отличия от родительских форм. Изменчивость вместе с наследственностью представляют собой два неразделимых свойства живых организмов.
Синдром Дауна

Синдром Дауна - генетическое заболевание, при котором кариотип состоит из 47 хромосом у человека вместо обычных 46. Это одна из форм анеуплоидии, о которой говорилось выше. В двадцать первой паре хромосом появляется добавочная, которая привносит лишнюю генетическую информацию в геном человека.
Название свое синдром получил в честь врача, Дона Дауна, который открыл и описал его в литературе как форму психического расстройства в 1866 году. Но генетическая подоплека была обнаружена почти на сто лет позже.
Эпидемиология

На данный момент кариотип в 47 хромосом у человека встречается один раз на тысячу новорожденных (ранее статистика была иной). Это стало возможным благодаря ранней диагностике данной патологии. Заболевание не зависит от расы, этнической принадлежности матери или ее социального положения. Оказывает влияние возраст. Шансы родить ребенка с синдромом Дауна возрастают после тридцати пяти лет, а после сорока соотношение здоровых детей к больным равняется уже 20 к 1. Возраст отца старше сорока лет также увеличивает шансы на рождение ребенка с анеуплоидией.
Формы синдрома Дауна
Наиболее частый вариант - появление дополнительной хромосомы в двадцать первой паре по ненаследственному пути. Он обусловлен тем, что во время мейоза эта пара не расходится по веретену деления. У пяти процентов заболевших наблюдается мозаицизм (дополнительная хромосома содержится не во всех клетках организма). Вместе они составляют девяносто пять процентов от общего количества человек с этой врожденной патологией. В остальных пяти процентах случаев синдром вызван наследственной трисомией двадцать первой хромосомы. Однако рождение двух детей с этим заболеванием в одной семье незначительно.
Клиника
Человека с синдромом Дауна можно узнать по характерным внешним признакам, вот некоторые из них:
Уплощенное лицо;
- укороченный череп (поперечный размер больше продольного);
- кожная складка на шее;
- складка кожи, которая прикрывает внутренний угол глаза;
- чрезмерная подвижность суставов;
- сниженный тонус мышц;
- уплощение затылка;
- короткие конечности и пальцы;
- развитие катаракты у детей старше восьми лет;
- аномалии развития зубов и твердого неба;
- врожденные пороки сердца;
- возможно наличие эпилептического синдрома;
- лейкозы.
Но однозначно поставить диагноз, основываясь только на внешних проявлениях, конечно, нельзя. Необходимо провести кариотипирование.
Заключение
Ген, геном, хромосома - кажется, что это просто слова, значение которых мы понимаем обобщенно и весьма отдаленно. Но на самом деле они сильно влияют на нашу жизнь и, изменяясь, заставляют меняться и нас. Человек умеет подстраиваться под обстоятельства, какими бы они ни оказались, и даже для людей с генетическими аномалиями всегда найдется время и место, где они будут незаменимы.