Mašina Golgi izvodi sljedeće funkcije. Struktura i funkcije GOLGI kompleksa. Lysosomes. Ćelijske inkluzije
Golgi kompleks To je snop membranskih torba (tenkova) i sistem mjehurića koji je s njim povezan.
Na vanjskoj, konkavnoj strani hrpa iz mjehurića koji su mjehurići od dragog. EPS, novi tenkovi se neprestano formiraju, a na unutrašnjoj strani spremnika pretvara se natrag u mjehuriće.
Glavna funkcija GOLGI kompleksa je prevoz tvari u citoplazmi i vanselularnoj mediju, kao i sintezu masti i ugljikohidrata. Golgji kompleks sudjeluje u rastu i ažuriranju plazmi membrane i u formiranju lizosoma.
Kompleks Golgi otvoren je 1898. godine K. Goldzhi. Imajući izuzetno primitivnu opremu i ograničen skup reagenata, učinio je otkriće, zahvaljujući kojem je Nobelovu nagradu primio s Ramon-I-Kahalom. Tretirao se nervnim ćelijama s otopinom bihromata, nakon čega su dodali nitrati srebra i osmija. Koristeći taloženje osmijske ili srebrne soli sa ćelijskim strukturama, tamno obojena mreža koja se nalazi u neuronima, koja se nazivala unutrašnjim mrežnim uređajima. Kada slikate opće metode, tanjurni kompleks ne akumulira boje, tako da je zona njene koncentracije vidljiva kao svijetlo područje. Na primjer, lagana zona je vidljiva u blizini jezgre plazme, koja odgovara području organizacije organela.
Najčešće, golgijski kompleks ide u kernel. S laganim mikroskopijom može se distribuirati u obliku složenih mreža ili pojedinačnih difuznih mjesta (dokumenata). Oblik i položaj organela nemaju temeljnu vrijednost i mogu se razlikovati ovisno o funkcionalnom stanju ćelije.
GOLGI kompleks je lokacija kondenzacije i akumulacije iz sekret proizvoda proizvedenih u drugim područjima ćelije, uglavnom u EPS-u. Tokom sinteze proteina, aminokiseline označene radioizotopom akumulira u gr. EPS, a zatim ih pronađite u GOLGI kompleksu, sekretornim uključivanjem ili lizosomima. Ovaj fenomen omogućava vam da odredite vrijednost Golgjie kompleksa u sintetičkim procesima u ćeliji.
Elektronska mikroskopija pokazuje da se GOLGI kompleks sastoji od klastera ravnih tenkova, koji se nazivaju Doniomas. Rezervoari su čvrsto susjedni jedni prema drugima na udaljenosti od 20 ... 25 nm. Čišćenje cisterna u središnjem dijelu od oko 25 Nm, a na periferiji, formiraju se ekspanzija - ampule, čija je širina neplatirala. U svakom snopu od oko 5 ... 10 tenkova. Pored čvrsto smještenih ravnih rezervoara u zoni GOLGJI kompleksa, postoji veliki broj malih mjehurića (vesicula), posebno na rubovima organela. Ponekad se deponuju iz ampula.
Sa strane pored EPS-a i kernela, nalazi se zona u Golgji kompleksu koja sadrži značajnu količinu malih mjehurića i malih tenkova.
Golgi kompleks je polariziran, odnosno visokokvalitetni je heterogeni sa različitih strana. Ima nezrelu površinu CIS-a koja leži bliže kernelu, a zrela - trans površina okrenuta prema površini ćelije. U skladu s tim, organela se sastoji od nekoliko međusobno povezanih odjeljaka koji vrše određene funkcije.
CIS pretinac obično se upućuje na Cell Center. Njegova vanjska površina ima konveksnu obrazac. Mikrovalne pećnice (transportnocitotički mjehurići), okrenut iz EPS-a, spojen sa tenkovima. Membrane se neprestano ažuriraju zbog mjehurića i zauzvrat ispunjavaju sadržaj membranskih formacija drugih odjeljaka. Odjeljak započinje obradu proteina nakon prijenosa, koja se nastavlja u sljedećim dijelovima kompleksa.
Međusobna operacija obavlja glikozilaciju, fosforilaciju, karboksilaciju, sulfatizaciju biopolimernih proteinskih kompleksa. Javlja se takozvana post-prevođenje modifikacija polipeptidnih lanaca. Postoji sinteza glikolipida i lipoproteina. U intermedijarnom komparatu formiraju se tercijarni i kvarterski proteinski kompleksi. Dio proteina podvrgnut je djelomičnoj proteolizi (uništavanje), što je popraćeno njihovom transformacijom potrebnom za zrenje. Stoga su CIS - i srednji pretinci potrebni za zrenje proteina i drugih složenih biopolimernih spojeva.
Trans-pretinac je bliži periferiji ćelije. Vanjska površina obično je konkavna. Djelomično transeg pretinca ulazi u Trans mrežu - sistem vezikula, vakula i tubula.
U ćelijama, pojedinačni dosadašnji praznici mogu biti povezani sa jednim drugim sistemom Vesiclea i tenkova, pored distalnog kraja akumulacije ravnih vrećica, tako da se formira labava trodimenzionalna mreža - Trans mreža.
Sortiranje proteina i drugih tvari, formiranje sekretornih granula, prethodnika primarnih lizosoma i spontanih sekretnih mjehurića javlja se u strukturama transporta i trans-mreže. Sekretorni mjehurići i predmiri surround proteini - klase.
Časovi se deponuju na membrani mjehurića koji se formiraju, postepeno se cijepi iz distalnog spremnika kompleksa. Furifikovani mjehurići odlaze iz Trans mreže, njihovo kretanje ovisi o hormonu i kontrolira funkcionalno stanje ćelije. Proces transporta mjehurića mjehurića je pod utjecajem mikrotubula. Proteini (Clalat) Kompleksi oko mjehurića raspada nakon cijepljenja mjehurića iz Trans mreže i ponovnog obrasca u vrijeme izlučivanja. U vrijeme izlučivanja, proteinski kompleksi mjehurića komuniciraju s MicroTubule proteinima, a mjehurić se prevozi u vanjsku membranu. Spontani sekretni mjehurići nisu okruženi nastavom, njihovo je formiranje kontinuirano i oni kreću prema staničnoj membrani, spajanje s njom, pružajući oporavak citlemme.
Općenito, Golgji kompleks sudjeluje u segregaciji - ovo razdvajanje, odvajanje određenih dijelova od rasuti i nakupljanje proizvoda sintetizirane u EPS-u, u njihovom prestrukturiranju hemikalija, sazrijevanje. U tenkovima se javlja sinteza polisaharida, njihova veza sa proteinima, što dovodi do formiranja složenih kompleksa peptidoglycana (glikoproteina). Uz pomoć elemenata GOLGI kompleksa, spremne tajne izvan granica sekretorne ćelije.
Mali transportni mjehurići su podijeljeni od gr. EPS u ribosomima Besplatne zone. Mjehurići vraćaju membrane kompleksa Golgi i isporučuju polimerne komplekse koji su sintetizirani u EPS-u. Mjehurići se prevoze u CIS-reklamu, gdje se spajaju sa svojim membranama. Stoga su novi dijelovi membrana i proizvoda sintetizirani u GR ubačeni u GOLGI kompleks. EPS.
U spremnicima kompleksa Golgi, na proteinima se pojavljuju sekundarne promjene u grčevima sintetizirane u gr. EPS. Ove promjene povezane su s restrukturiranjem lanaca od glikoproteina oligosaharide. Unutar šupljina GOLGJI kompleksa s translukozidazama, lizosomalni proteini i proteini tajni su modificirani: dosljedna zamjena i produženje lanaca oligosahackeride. Modificiranje proteina prenose se iz cisterna CIS odjeljka u rezervoar preko pregrada zbog transporta u mjehurićima koji sadrže protein.
U trans-pretincu su proteini sortirani: proteinski receptori nalaze se na unutrašnjim površinama membrana spremnika, koje prepoznaju sekretarnje proteine, membrane i lizosome (hidrolaza). Kao rezultat toga, tri vrste finih vakuela cijene se iz distalnih trans-presjeka: koji sadrže hidrolazu - pre-seanse; Sa tajnim jedinicama, vakuole koje nadopunjuju staničnu membranu.
Sekretna funkcija kompleksa Golgji je da se izvezeni protein sintetizira na ribosomima, odvojenim i akumulirajući unutar spremnika EPS-a, prevozi se u vakuolu ploče uređaja. Tada se akumulirani protein može kondenzirati, formirati sekretarnje granule proteina (u pankreatičnoj, laktičkoj i drugoj žlijezdi) ili ostaju raspušteni (imunoglobulini u plazmi). Od ampularnih ekstenzija rezervoara u golgi kompleksu, mjehurići koji sadrže ove proteine \u200b\u200bsu cijepljeni. Takvi se mjehurići mogu spojiti među sobom, povećavati veličinu, formiranje sekretarnih granula.
Nakon toga, sekretarne granule počinju prelazeći na stanicu, u kontaktu sa plazmom, sa kojim se spajaju vlastiti membrane, a sadržaj granula ispadaju izvan ćelije. Morfološki, ovaj se proces naziva ekstruzija ili izlučivanje (pražnjenje, exocitoza) i podseća na endocitozu, samo obrnutom redoslijedom faza.
GOLGIE kompleks može se oštro povećavati veličine u ćelijama koje aktivno obavljaju sekretornu funkciju, koja obično prati razvoj EPS-a, a u slučaju sinteze proteina, nukleina.
Tijekom podjele ćelije, složeni Golges razdvajaju pojedine tenkove (DiSokooma) i / ili mjehuriće, koji se distribuiraju između dvije dijeljenje i na kraju slona vraćaju strukturni integritet organele. Iz odjela se nalazi kontinuirano ažuriranje membranske aparata zbog mjehurića koji migriraju iz EPS-a i distalnih prepozova zbog proksimalnih odjeljaka.
Ako ste našli grešku, odaberite fragment teksta i kliknite Ctrl + Enter..
Struktura i funkcije GOLGI kompleksa povezane su s popunjavanjem izmjene tvari koje dolaze iz EPA-e i njihove preraspodjele na svoje odredišne \u200b\u200bstavke.
U životinjskim ćelijama, najčešće je jedan veliki set Golgija, u povrće - nekoliko manjih hrpa koji se nazivaju Doniomas.
U pogledu njegove strukture, Golgi aparat je snop membranskih diskova (sa šupljinama iznutra). Svaki takav disk se naziva tenk. Svaki tenk se širi na ivice. Pored diskova, uključeni su i vezivnici povezani sa njima, kao i (vjerojatno) okolna membrana mreža koja povezuje pojedinačne tenkove zajedno.
Golgina strana okrenuta je kernelu naziva se CIS-om. Strana okrenuta plasmamemme je pretoska dubina. Takođe izdvojite srednju odeljenje. Enzimski sastav raznih odjela se izliva, dakle, u svakom od njih se pojavljuju hemijske reakcije, to su faze modifikacije tvari. Supstanca koja prolazi kroz tenkove kao u transporteru, postepeno stječe potrebne hemijska struktura i funkcionalnost.
Iz endoplazmatske mreže, proteini, masti i ugljikohidrate sintetiraju se tamo u Golgji kompleksu uz pomoć vize (mjehurići okruženi membranom). Istovremeno, proteini imaju signalne hemijske oznake (u obliku oligosaharida), koji "prijavljuju" kompleks Golgji, šta da radim sa njima.
U ovom obrascu dijagram prikazuje kao protein koji je sintetiziran u EPS, prolazeći kroz Golgi aparat, postaje komponenta ćelijske membrane. Proteini je ovdje označen zelenim ovalnom. Ružičasti element priključen na njega ukazuje na ugljikohidrat povezan sa proteinom. U suštini se prevozi i modificirani ne proteini, ali glikoprotein (ugljeni hidrat + protein).
Izgradnja citoplazmatske membrane samo je jedna od funkcija GOLGI kompleksa. Također izvan ćelija egzocitozom, komponente međućelijske tečnosti, matrica ćelijskih zidova (u biljkama), razne tajne (u sekretarnim ćelijama) itd.
Druga je funkcija formiranje lizosoma - mobilne organele koje sadrže uglavnom enzime za raspad u dolaznim složenim tvarima.
Također u golubama se formiraju u transportne vezike, pružajući tvari na druge stanične organele.
Kompleks ili aparat, Golgi se naziva tako u čast naučnika otkrivenog. Ova organa za ćeliju ima oblik skupa šupljina ograničen na pojedinačne membrane. U biljnim ćelijama i najjednostavnijim su prikazani nekoliko zasebnih manjih hrpa (DontIOMAS).
GOLGI kompleks izgleda naizgled u elektronskom mikroskopu, podseća na hrpu vrećica u obliku diskoteka koji se nalaze jedna na drugu, o kojima postoji mnogo mjehurića. Unutar svake "vrećice" je uski kanal, širenje na krajevima u takozvanim tenkovima (ponekad se tenk naziva cijela torba). Mjehurići su mjehurići od njih. Oko centralnog snopa formiran je sistem međusobno povezanih cevi.
Sa vanjskom, imajući ne pomalo konveksne formi, stranice hrpa formiraju novi tenkovi spajajući mjehuriće obračun iz glatke endoplazmatske mreže. Na unutrašnjoj strani rezervoara, rezervoari se završavaju i ponovo raspadaju na mjehuriće. Dakle, tenkovi (hrpe hrpe) Golgji prelaze s vanjske strane do unutarnjeg.
Dio kompleksa, smješten bliže kernelu, naziva se "CIS".
Ona je bliža membrani, "trans".
Funkcije kompleksa Golgi
Funkcije Golgi uređaja su raznolike, ukupno smanjeno za izmjenu, redistribucija tvari sintetizirane u ćeliji, kao i njihovo uklanjanje ćelije, formiranje lizosoma i izgradnju citoplazmatske membrane.
Aktivnost GOLGI kompleksa je visoka u sekretornim ćelijama. Proteini koji dolaze iz EPS-a koncentrirani su u Golgi uređaju, a zatim prebačeni u membranu u Golgi Bubbles. Enzimi se izlučuju iz ćelije obrnutim pinocitozom.
Priloženi su lanci oligosahardida koji dolaze u Golgji. U aparatu su izmijenjeni i služe markeri, sa kojima su proteini sortirani i poslani na svoj način.
U biljkama, tokom formiranja ćelijskog zida Golgi, ugljikohidrati koji služe matricu za njega (celuloza ovdje nije sintetizirana). Burgrid Bubbles of Golges Pomicanje mikrotubula. Njihove membrane spajaju se sa citoplazmatskim membranom, a sadržaj su uključeni u stanični zid.

Golges složen kompleks (u debiliji epitelijuima crijevne sluznice i respiratornog trakta) izlučuje glikoprotein Muzina koji u rešenjima formira sluz. Takve supstance sintetizirane ćelije korijenskog vrha, lišća itd.
U ćelijama tankog crijeva, Golgi aparat obavlja funkciju lipidnog transporta. U ćelijama pada masne kiseline i glicerol. U glatkom EPIS-u dolazi do sinteze njihovih lipida. Većina ih je obložena proteinima i prevoze se u ćelijsku membranu. Prolazeći kroz to, lipidi su u limfi.
Važna funkcija je formiranje lizosoma.
Golgi kompleks - Ovo je membranska struktura svojstvena bilo kojoj eukariotskoj ćeliji.
Uređaji za golgi predstavljeni su spljoštenim tenkovima (ili vrećama) sakupljenim u snopu. Svaki rezervoar je malo zakrivljen i ima konveksnu i konkavnu površinu.
Prosječni promjer rezervoara je oko 1 μm. U središtu tenka njegove membrane su približne, a na periferiji često čine proširenja ili ampule, iz kojih su mjehurići povoljni. Paketi ravnih cisterna sa prosjekom od oko 5-10 formiranih docyoma. Uz tenkove, u kompleksu Golgja prisutni su transportni i sekretorirani mjehurići. U DonteoMomeu, u skladu sa smjerom zakrivljenosti zakrivljenih površina, tenkovi se odlikuju dvije površine. Konveksna površina naziva se nezrela ili površina CIS-a. Adrešen je kernelu ili na tubularni endoplazmatski mrežni kanali i povezan je s posljednjim mjehurićima, odstupio iz zrnačke mreže i dovedi molekulu proteina u docyma na zrenje i dizajn u membrani. Suprotni transiosiosomi su konkavni. Suočen je sa plasmolemom i naziva se zrelom jer se iz njegove membrane uzimaju mjere za sekretne mjehuriće koji sadrže gotove proizvode izlučivanja.
Golgji kompleks sudjeluje:
- u akumuliranju proizvoda sintetizirane u endoplazmatskoj mreži,
- u njihovom hemijskom restrukturiranju i sazrijevanju.
U tenkovima kompleksa Golgi, dolazi do sinteze polisaharida, njihova složena sa molekulama proteina.
Jedna od glavnih funkcija Golgjie kompleksa je formiranje gotovih sekretornih proizvoda koji su izloženi izvan ćelija egzocitozom. Najvažnije funkcije kompleksa Golgji su takođe ažuriranje ćelijskih membrana, uključujući presjeke PLASMOLM-a, kao i zamjenu plazmolm oštećenja u procesu sekretarnih aktivnosti ćelija.
Golges Complex smatra se izvorom formiranja primarnih lizosoma, iako su njihovi enzimi sintetizirani u zrnatu mrežu. Lisosomi su intrarealno u nastajanju sekretarskih vakutivnih vakulacija napunjenih hidrolitičkim enzimima potrebnim za procese fago i autofagocitoze. Na laganom optičkom nivou, lizosomi se mogu tumačiti i suditi o stupnju svog razvoja u ćeliji u aktivnosti histohemijske reakcije na kiselo fosfatazu - ključni lizosomalni enzim. U elektronskoj mikroskopiji lizosomi su definirani kao mjehurići ograničeni hiloplazmom membrane. Uvjetno raspoređivanje 4 glavne vrste lizosoma:
- primarni
- sekundarni lizosomi,
- autofagosomi
- preostali Bik.
Primarni lizosomi su male membranske mjehuriće (prosječni promjer je oko 100 nm) ispunjen homogenim finim sadržajima, koji je skup hidrolitičkih enzima. Oko 40 enzima (protabi, nukleasti, glikozidaze, fosforidaze, sulfataze) identificiraju se u Lysosomima, optimalni način djelovanja dizajniran je za kiseli medij (pH 5). Lizosomalne membrane sadrže posebne prevozne proteine \u200b\u200bza prevoz od lizosoma u hiloplazmima hidroličnih cerišća - aminokiselina, šećera i nukleotida. Lesos membrana otporna je na hidroličke enzime.
Sekundarni Lysosomi formiraju se kada su primarni lizosomi prekriveni endocitozom ili pinocitnim vakuolima. Drugim riječima, sekundarni lizosi su unutarćelijski probavni vakuola, čiji su enzimi isporučuju primarni lizosomi i materijal za probavu - vakuol endocitoze (pincite). Struktura sekundarnih lizosoma vrlo je raznolika i varira u procesu hidroličnog dijela sadržaja. Enzimi Lizosoma podijelila je biološke tvari u ćeliju, rezultirajući monomeri, koji se prevoze kroz lesosoma membranu u hiloplazmi, gdje se koriste ili uključuju u različite sintetičke i metaboličke reakcije.
Ako interakcija s primarnim lizosomima i hidroliznim dijeljenjem njihovih enzima podliježe vlastitim ćelijskim strukturama (starenje orgele, uključivanjem itd.), Formirana je autofagomija. Autofagocitoza je prirodni proces u vitalnim ćelijama ćelije i igra veliku ulogu u ažuriranju svojih građevina sa intracelularnom regeneracijom.
Preostali Bik Ovo je jedna od završnih faza postojanja fago-autizma i nalaze se u nepotpunoj fazi ili autofagocitozi i nakon toga pušta iz ćelije egzocitozom. Oni su zbijali sadržaj, često se promatra sekundarno strukturiranje neiskorištenih spojeva (na primjer, lipidi tvore složene slojevljene formacije).
Društveni gumbi za Joomla
Funkcije kompleksa Golgi
1. Sinteza polisaharida i glikoproteina (glikocalix, sluz).
2. Obrada molekula:
a) terminalni glikozilacija
b) fosforilacija
c) sulfatizacija
d) proteolitički dijeljenje (dijelovi molekula proteina)
3. Kondenzacija sekretornog proizvoda.
4. Pakovanje sekretornog proizvoda
5. Razvrstavanje proteina u zoni Trans-Jolly mreže (zbog određenih membranskih proteina receptora koji prepoznaju područja signala na makromolekulama i usmjeravaju ih u odgovarajuće mjehuriće). Prevoz iz kompleksa Golgi ide u obliku 3 potoka:
1. Hidrolazni mjehurići (ili primarni lizosomi)
2. U plazmilu (u sastavu mjehurića)
3. U sekretarnim granulama
Endosomi - Membranske mjehuriće s plačenim sadržajem i pružanje molekula u ćeliji. Vrsta prijenosa tvari endos sustava različita:
1. Sa probavom makromolekula (punih)
S djelomičnim dijeljenjem
3. Bez promjene u toku prevoza
Proces transporta i naknadno oduzimanje tvari u ćeliji pomoću endosome sastoji se od sljedećih uzastopnih komponenti:
1. rano (periferno) endosome
2. kasno(perinucle) endosome predvostručena faza probave
3. lizosoma
Rani endosome - lišen mjehurića Clatrine na periferiji ćelija. PH od 6,0 \u200b\u200bokoliša, ovdje je ograničen i podesiv proces cijepanja (ligand je odvojen od receptora) - povrat receptora u ćelijsku membranu. Rano endosome je i dalje poznat kao curl.
Kasno (perinuklearno) endosome: a) kiseli kiseli sadržaj pH 5.5
b) Prečnik veći do 800 nm
c) dublji nivo probave
Ovo je probava liganda (periferni endoseme + perinuklearni endosome) - Višetaurički pozivaoca.
Lysosomes
1. Fagolizosoma - Formira se kada je zbrka kasnih endosoma ili lizosoma sa fagozomom. Proces uništavanja ovog materijala naziva se heterofage.
2.Autofagolizosoma- Formira se kada je zbrka kasnih endosoma ili lizosoma sa autofagosomom.
3. Mulvezikularne priče- Veliki vakuola (800 Nm), koji se sastoji od malih mjehurića od 40-80 Nm okružena umjereno gustom matricom. Formira se kao rezultat spajanja ranih i kasnih endosa.
4. Preostale priče- Ovo je neimeizvani materijal. Najpoznatija komponenta ove vrste je lipofUscin granule - mjehurići dia. 0,3 - 3 mikrona koji sadrže lipofuscinski pigment.
Cytoskeletton - Ovo je sistem mikrotubula, mikrofilamenta (srednji, mikrotracekal). Svi oni formiraju trodimenzionalnu mrežu, interakciju sa mrežama iz drugih komponenti.
1. Mikrotubule- Šuplji cilindri dia. 24-25 Nm, debljina zida 5 nm, dia. Lumen je 14-15 Nm. Zid se sastoji od spiralno postavljenih niti (nazivaju se protofilamentima) debljine 5 nm. Ove niti formiraju dimeri i tubulin. Ovo je labilni sistem, koji ima jedan kraj (naznačeno "__") je fiksiran, a drugi ("+") je besplatan i sudjeluje u procesu depolimerizacije.
MicroTubule.povezana sa brojnim proteinima koji imaju opšte ime ožu - oni vežu mikrotubule sa drugim elementima citoskeleta i organela. Kinesin - (korak njegovog kretanja duž površine mikrotubula je 8 Nm).
Orgela
sl. MicroTubule
Mikrofilamenti- Ovo su dva upletena F-djela za koje su sastavljene od G-Actin-a. Promjer njih je 6 nm. Mikrofilamenti Polar, dodavanje G -kortina javlja se na ("+") kraj. Oni formiraju klastere
prema periferiji ćelija i povezani su sa plazmom kroz srednje proteine \u200b\u200b(-aktin, vinculin, talin).
Funkcija: 1. Promijenite citosol (prijelaz Sol u gel i nazad).
2. Endocitoza i exocitoza.
3. Mobilnost ćelija koji nisu ježevi.
4. Stabilizacija lokalnih dosega plazme membrane.
Intermedijarna nitimaju d 8-11 nm, sastoji se od proteina karakterističnih određenih tipova ćelija. Oni čine intracelularni okvir koji pruža elastičnost ćelije i naručenu lokaciju komponenti citoplazma. Srednji filamenti formiraju se molekulama proteina sličnim filamentima, utkani jedni s drugima poput konopa.
Funkcije: 1. Strukturni
2. Učešće u formiranju roga
3. Održavanje oblika, nervnih procesa
4. Pričvršćivanje miofibrila na plasmolem.
Mikrotraboni- Otvorite mrežu tankih pređe, koji postoje u kompleksu s mikrotubulima i može učestvovati u prevozu organele i utjecati na viskoznost citosola.
Predavanje
Tema: "Core. Struktura interfaze kernela. Osnove biosintetičke stanice "
Jezgro Glavni je dio ćelije kodiranja informacija o strukturi i funkciji organa. Te se informacije postavljaju u genetski materijal, DNK, koji je u kompleksu s osnovnim proteinima (histone) DNP-a. Za neku iznimku (mitohondrija), DNK se lokalizira isključivo u kernelu. DNK se može ponoviti, čime se osigurava prijenos genetičkog kodeksa dječijim ćelijama u ćeliji.
Kernel igra središnju ulogu u sintezi proteina i polipeptida, što je nosilac genetskih informacija. Sva jezgra ćelijskog ćelija sadrže iste gene, neke ćelije su različite u svojoj strukturi, funkcijama i karakteru proizvedenih tvari. Nuklearna kontrola vrši
represije ili depresija (izraz) aktivnosti različitih gena. Emitovanje sinteze proteina povezano je sa formiranjem M-RNA. Mnogi RNA su kompleks proteina i rna, tj. RNP. Interfase jezgra u većini ćelija je formiranje zaobljenog ili ovalnog oblika u promjeru nekoliko mm. U leukocitima i ćelijama vezivnog tkiva, kernel je unosan i naznačen term polimorfnim.
Interfase kernelima nekoliko različitih struktura: nuklearna školjka, kromatin, kariolimf i nukleolo.
Nuklearna školjka
1. vanjska nuklearna membrana- Ribosomi se nalaze na površini, gdje su proteini ulaze u periklearni tenkove sintetizirane. Sa citoplazme, okružen je labavom mrežom srednjih (vimenzin) filamenata.
2. Perinuklearni rezervoari- Dio obrubnih rezervoara povezan je s granulovnim endoplazmatskim mrežama (20-50 Nm).
3. Interna nuklearna membrana - odvojen od sadržaja kernela s nuklearnom pločom.
4. Nuklearna pločadebljina 80-300 Nm, sudjeluje u organizaciji nuklearne školjke i perinuklearnog kromatina, sadrži proteine \u200b\u200bsrednjih filamenata - laminati A, B i C.
5. Nuklearno vrijeme - od 3-4 hiljade specijaliziranih komunikacija, prevoza između jezgre i citoplazme. Nuklearno vrijeme D 80 NM, ima: a) pore kanal - 9 nm
b) Potonje sadrži proteina receptora koji odgovara na nuklearni uvoz (ulazna karta do kernela). Nuklearni pore može povećati promjer poreze i prebacivanje na jezgro velikih makromolekula (DNK RNA polimeraza) .
Nuklearno vrijemesastoji se od 2 paralelnih prstenova jedan po jedan sa svakom površinom cariolema. Prsten s promjerom od 80 Nm, formiraju ih 8 proteinskih granula, od svakog granula do centra, navoj (5 Nm) dostiže, koji tvori particiju (otvor). Centar je središnja granula. Kombinacija ovih struktura se zove nuklearni pore kompleks.Cannel s promjerom od 9 Nm generira se ovdje, takav se kanal naziva voda, jer se mali molekuli i joni vode u vodi se kreću duž.
Funkcije nuklearne pore: 1. izborni prijevoz;
2. Aktivni prenos u kernel proteina s nizom karakteristika proteina nuklearne lokalizacije;
3. Prebacite na citoplazmu ribosoma subitineta s promjenom u skladu sa pore kompleksom.
Unutrašnja nuklearna membrana- Glatko i povezano uz pomoć integralnih proteina nuklearnom pločom, koja je sloj, debljina 80-300 nm. Ova ploča ili lamin- Sastoji se od isprepletenih srednjih filamenata (10 nm) koji formiraju karyarek. Njene funkcije:
1. Spremanje strukturna organizacija Pore \u200b\u200bkompleksi;
2. održavanje oblika kernela;
3. Naručeni ležeći kromatin.
Formira se kao rezultat spontanog udruženja 3 glavne polipeptide. Ovo je strukturalni okvir nuklearne školjke sa dijelovima specifičnog vezivanja kromatina.
Stroj Golgi.
Poglavlje 1. Mašina Golgji: Struktura i funkcije
Stroj Golgi.
1.1. Golgi Auto: Struktura
Opis strukture Golgi uređaja usko je povezan s opisom osnovnih biohemijskih funkcija, jer se podjela ETCO-ćelijskog odjela na odjeljenjima vrši uglavnom na osnovu lokalizacije enzima ...
Stroj Golgi.
1.2. Golgi Auto: Funkcije
Funkcija Golgi uređaja je transportna i hemijska izmjena tvari koje ulaze u njega. Početna supstrata za enzime su proteini koji ulaze u uređaj Golgja iz endoplazmatskog retikuluma ...
Stroj Golgi.
Poglavlje 2. Analiza aktivnosti mašina u ćeliji
Stroj Golgi.
2.1. Analiza aktivnosti strojeva Golgi u ćeliji
Lizosomi su mali, okruženi jednoj membranskim mjehurićima. Oni su pupljeni iz Golgi aparata i možda, iz endoplazmatskog retikuluma. Lizosomi sadrže razne enzime koji podijeli velike molekule ...
Stroj Golgi.
2.3. Golgi aparati: vrsta proteina i prijenosa signala
Golges kompleks djeluje na raskrižju sekretornih staza, provodeći prijem novo sintetiziranih proteina i lipida iz ER, njihove kovalentne modifikacije, a zatim - sortiranje reakcijskih proizvoda u skladu s njihovom svrhom (Sl. 1 gg) ...
Stroj Golgi.
2.3. Strojevi: Molekularni mehanizam rada
Heptamer Cytosol protein kompleks, zvani Copi (membratni kompleks, Boomer), u vezi sa GTP-obvezujućim proteinskim ARF-om 1 formira školjku na takav način da se pridruži u golgi membrani ...
Genettawic
1.MeeTecraft
Uređaji sa sastoji se od mokraćnih organa koji osiguravaju formiranje i uklanjanje mokraćnih i genitalnih organizmi koji provode funkciju reprodukcije. Funkcionalno, nisu povezani jedni s drugima ...
Značajke strukture ptica
Aparati za probavu
Struktura probavnog sustava ptica u velikoj mjeri podsjeća na sisarnog probavnog aparata. Uključuje fliplot, jednjak i želucu, tanke i debele crijeve. Priroda prerade čvrstog hrane ...
Značajke strukture ptica
Mašina disanja
Respiratorni organi u pticama imaju brojne karakteristike: nisku vrijednost i jednostavnost strukture nosalne šupljine; Prisutnost u bifurciranju trake uređaja za emitiranje zvuka - pjevači larinx; Manja veličina i položaj pluća ...
Značajke strukture ptica
Uređaji za mokraće
U mokraćnog sistema sastoji se samo od otvaranja bubrega i uremara u waggery of Cloca.
Lohanka, mjehur, mokraćni kanali u pticama nedostaju ...
Značajke strukture ptica
Uređaji za reprodukciju
Sistem reproduktivnih tijela osigurava nastavak nagađanja. U poljoprivrednim pticama također određuje proizvodnju jaja. Ovaj sistem se sastoji od spolnih žlijezda (sjemenki ili jajnika) u kojima se formiraju seks ...
Uloga vizuelnog analizatora u životu životinja
1.4 Kontinski aparat
Oko se može posmatrati kao optička kamera. Da biste vodili takvu "kameru" na objektu pod pregledanjem (tačka fiksacije), treba ga rotirati. Za pokrete očne jabučice postoje čaše ...
Fotodinamički efekat i fotodinamička terapija
10. Golgi i endoplazmatski reticulum aparat
Hidrofobični fotonsizeri, poput hipericina, PC 4 Phtalocijanin, pocinčan fhalocijanin ili photofrine, obično se akumuliraju u perinuklearnom području bogatom membranim organelima - mitohondria, eteri ...
Čestovni europski dio Rusije sa dnevnim efektima života
3.1.1 VOZILO ROTA
Usmeni aparat struganja nastale su specijalizacijom običnih udova artropoda. Apsorpcija i mljevenje hrane. Usta leptira jednako su karakteristična karakteristika od strukture krila i pokrivaju svoje vage ...
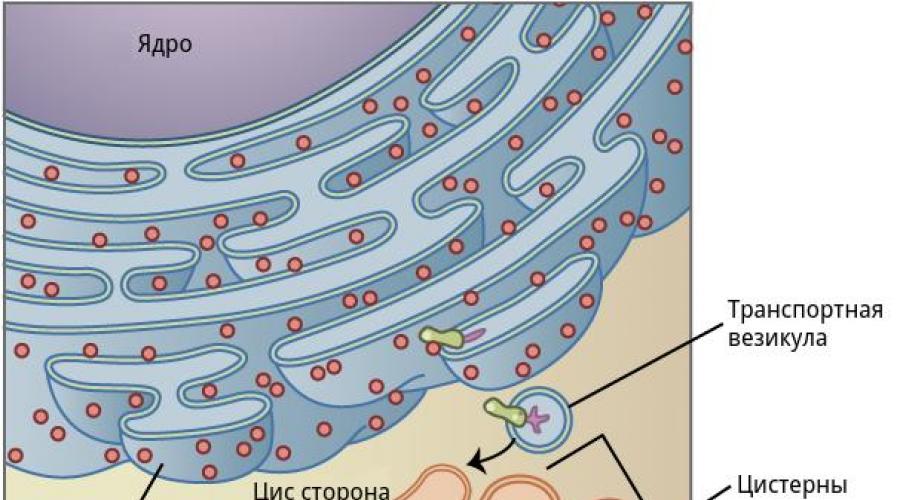
GOLGI kompleks je snop membranskih vrećica (tenkova), lagano proširen bliže rubovima i pridruženi sistem golgih mjehurića.
Gotovo sve izlučene ćelije supstancije (i proteine \u200b\u200bi neetinske prirode) prolaze kroz strojeve i postoje pakirane u sekretornim mjehurićima. Membranski elementi sudjeluju u segregaciji i akumuliranju proizvoda sintetizirane u ER, sudjeluju u njihovom prestrukturiranju hemikalija, zreći: to je uglavnom restrukturiranje oligosahardidnih komponenti glikoproteina u sastavu tajni topljivih voda ili kao dio membrane.
U tenkovima se javlja sinteza polisaharida, njihov odnos sa proteinima, što dovodi do formiranja sluznica. Ali najvažnije, uz pomoć elemenata Golgi aparata, proces uklanjanja gotovih tajna izvan ćelije. Pored toga, AG je izvor ćelijskih lizosoma.
Učešće AG-a u procesima uklanjanja sekretornih proizvoda bilo je vrlo dobro proučano primjerom explokrilne stanice pankreasa. Za ove ćelije prisustvo velikog broja sekretornih granula (zimske granule), koje su membranske mjehuriće ispunjene sa sadržajem proteina. Kao dio proteina zimskih granula uključuje razne enzime: protabi, lipaze, ugljikohidraze, nukleaste. Kada se izlučuju sadržaj ovih zimskih zrnca izbacuje se iz ćelija u klirensu, a zatim teče u šupljinu creva. Budući da je glavni proizvod ćelija gušterače ćelija proteina, istraženo je redoslijed uključivanja radioaktivnih aminokiselina u raznim ćelijskim odjeljcima. U tu svrhu, životinje su ubrizgane s vrstom aminokiseline (3 N-leucina) i uz pomoć elektrona mikroskopskih autograma nadgleda se preko lokalizacije etikete. Pokazalo se da je nakon kratkog vremenskog perioda (3-5 min), naljepnica bila lokalizirana samo u bazalnim područjima ćelija, u parcelu bogata granularnoj. Budući da je naljepnica bila uključena u proteinski lanac tokom sinteze proteina, bilo je jasno da niti se niti sinteza proteina ne pojavljuje u niti u zimskim granulama, a ona se sintetizira isključivo u ergastoplazmu na ribosomima. Nešto kasnije (nakon 20-40 minuta) na etiketi osim Ergastoplasma nalazi se u zoni Vakolus AG. Stoga, nakon sinteze u Ergastoplazmi, protein je prevezen u AG zonu. Kasnije (nakon 60 minuta) naljepnica je pronađena već u zoni zimskih granula. Ubuduće se naljepnica mogla vidjeti u aspektu acinusa ove žlijezde. Stoga je postalo jasno da je Ah intermedijarna veza između sinteze same izlučenog proteina i eliminacije od njega iz ćelije. Također detaljno o procesima sinteze i uklanjanja proteina proučavani su na drugim ćelijama (mlijeko gvožđe, zidane crijevne ćelije, štitne žlijezde itd.) I morfološke karakteristike ovog procesa. Izvezeni proteini sintetizirani na ribosomima odvojen je i akumuliran unutar er tenkova duž kojih se prevozi u membran. Ovdje su mali vakuole koji sadrže sintetizirane proteine \u200b\u200bkoji dolaze u zonu vakula u proksimalnom dijelu DontIOMMOM-a, ovdje se cijene iz glatkih dijelova ER-a. Na ovom mjestu, vakuole se mogu spojiti jedni s drugima i sa ravnim ciferencijama neposvojenog. Stoga se proteinski proizvod prenosi već unutar šupljina hipertenzije AG.
Kako su proteini modificirani u cisternima Golgi uređaja, oni se prebacuju iz malih vakula iz spremnika do tenka do distalnog dijela Dontyoma, sve dok ne dođu do tubularne membrane mreže u dontiosnom trans. U ovom se području čisti cijepanje malih mjehurića koji sadrži zreli proizvod. Citoplazminalna površina takvih mjehurića slična je površini mjehurića mjehurića, koji se promatraju s pinocitozom receptora. Odvojeni fini mjehurići spajaju se jedno s drugim, formirajući sekretne vakuleve. Nakon toga, sekretni vakuole počinju prelaze na površinu ćelije, dođu u kontakt sa plazma membranom, sa kojom se njihove membrane spajaju, a samim tim, sadržaj tih vakula je izvan ćelije. Morphološki, ovaj proces ekstruzije (pražnjenja) podseća na pinocitozu, samo obrnutom redoslijedom faza. On je zvao exocitosis.
Takav opis događaja je samo opća šema za sudjelovanje Golgi aparata u sekretornim procesima. Slučaj je kompliciran činjenicom da ista ćelija može sudjelovati u sintezi mnogih proteina izdvojenih, mogu ih izolirati jedni od drugih i izradati na površinu ćelije ili na sastav lizosoma ili na sastav lizosoma. U Golgi uređaju, to nije samo "pumpanje" proizvoda iz jedne šupljine do druge, već i postepeno njihovo "sazrijevanje", modifikacija proteina, koji se završava "sortiranjem" proizvoda ili na lisosomam ili na lizosomam ili plazma membranaili na sekretna vakula.
Ulaznica 36.Modifikacija proteina u Golgi uređaju. Poredaj proteine \u200b\u200bu AG
U rezervoarima od Golgi, proteini su namijenjeni za sekreciju, transmembran plazma proteine \u200b\u200bmembrane, proteine \u200b\u200blizosoma itd. Zrukinja sukcesivno se kreću po spremnicima organela, u kojima su njihove izmjene modificirane - glikozilacije i fosforizacije. Kada su O-glikozilacije, složeni šećeri kroz kiseonik atoma na proteinima. U fosforizaciji je ostatak ortofosforične kiseline priložen za proteine. Različiti rezervoari u Golgi uređaju sadrže različite rezidencijalne katalitičke enzime i, stoga se u njima javljaju različiti procesi sa zrelim proteinima u njima. Jasno je da bi takav korak proces trebao nekako kontrolirati. Zaista, proteini sazrijevanja su "označeni" sa posebnim ostacima polisaharida (uglavnom mannose), očigledno igrajući ulogu svojevrsne "kvalitetne marke". Postoje dva hipoteza za međusobno povezanost koji objašnjavaju ovaj mehanizam:
· Prema prvom, prevoz proteina vrši se koristeći iste vezivne mehanizme iz veznog prevoza, kao i put prevoza iz EPR-a, a rezidencijalni proteini nisu uključeni u vezike za vezanje;
· Prema drugoj, neprekidno kretanje (sazrijevanje) samih tenkova, njihova skupština mjehurića s jednog kraja i demontaže s drugog kraja organela, i rezidencijalni proteini pomiču se retrogradno (u suprotnom smjeru) pomoću vezicular transporta.
| Poznato je da samo proteini prethodnici lizosoma hidrolize imaju specifičan oligosaharid, naime mannose grupu. U CIS-tenksima su ove skupine fosforirane i dalje zajedno s drugim proteinima prenose se iz tenka u tenk, kroz srednju zonu na trans-mjestu. Trans-mrežne membrane uređaja Golgji sadrže transmembranske proteine \u200b\u200b- receptor (mannose-6-fosfatni receptor ili M-6-F-receptor), koji učini fosforiklirane manalne grupe ligosahachide lanca lizosomalnih enzima i povezano je s njima . Ovo vezivanje se događa kod neutralnih vrijednosti pH unutar rezervoara preko mreže. Na membranama, ovi proteini M-6-F-F-receptora oblikuju klastere, grupe koje su koncentrirane u zonama formiranja malih mjehurića prekrivenih Clawninom. U preko mreži Golgi aparata su odvojeni, pupčani i daljnji prenos na endosome. Shodno tome, M-6-F-receptori, koji su prenosni proteini, obvezujući na Lysosom Hydrolause, razvrstani su, sortirani, od ostalih proteina (na primjer, sekretorni, nelioosomalni) i koncentriraju ih u mjehurićima. Skidanje od Trans mreže Ovi mjehurići brzo gube krzneni kaput, spajanje sa endosomima, prenoseći svoje lizosoma enzime povezane sa membranskim receptorima, na ovaj vakul. Kao što je već spomenuto, unutar endose zbog aktivnosti protonskog nosača, medij je postignut. Počevši od pH 6, lizosomalni enzimi koji se distribuiraju iz M-6-F-F-F-receptora, aktivira se i počnite raditi u šupljini endolize. Sekcije membrana zajedno sa M-6-F-receptorima vraćaju se recikliranjem membrane mjenjača natrag na preko mreže Golgi uređaja. Najvjerovatnije, taj dio proteina koji se nakupljaju u sekretarnim vakuletama i izveden je iz ćelije nakon što signal stigne (na primjer, nervni ili hormonalni) prenosi isti postupak odabira, sortiranje na receptoru za trans-rezervoar. Ovi sekretarnji proteini prvi se pojavljuju u malim vakulama obučene i sa Clawninom, koji se zatim spajaju jedno s drugim. U sekretarnim vakuletima, agregiranje akumuliranih proteina često se javlja u obliku gustih sekretarnih granula. To dovodi do povećanja koncentracije proteina u tim vakuelima u oko 200 puta, u poređenju sa koncentracijom u Golgi uređaju. Tada su ovi proteini, kako se nakupljaju u sekretornim vakuletima, izbačeni iz ćelije egzocitozom, poljem dobivanja ćelije odgovarajućeg signala. Treći protok vakuela povezanih sa konstantnom, konstitutivnim izlučivanjem dolazi iz Golgi aparata. Dakle, fibroblasti izdvajaju veliki broj glikoproteina i sluznica uključenih u glavnu supstancu vezivnog tkiva. Mnoge ćelije neprestano oslobađaju proteine \u200b\u200bkoji doprinose vezivanju njih sa podlozima, protok membranskih mjehurića neprestano je u toku elementovima glikoproteina za površinski nosač ćelija i membranske glikoproteine. Ovaj tok komponenti koje se oslobađa ćelijom ne podliježe sortiranju u prenosu receptora u Golgi uređaju. Primarni vakuole ovog toka također su cijepljene iz membrane i pripadaju njihovoj strukturi do ograničenih vakula koji sadrže Clatrine. Završavanje razmatranja strukture i rada takvih složenih membranskih organela, kao Golgi aparat, potrebno je naglasiti da uprkos naizgled morfološkog homogenosti njegovih komponenti, vakuola i tenkova, u stvari, to nije samo priobljivost mjehurića, Ali vitki, dinamični kompleks, organiziran, polarizirani sistem. U AG, ne samo prijevoz nije posetio od EP-a do plazme membrane. Postoji retrogradni prijenos Vesicula. Dakle, vakuole se cijene iz srednjih lizosoma i vraćaju se zajedno sa proteinima receptora u trans-AG zoni. Pored toga, postoji protok vakuola iz područja trans-zone do ZON-Zone AG, kao i iz ZO-zona do endoplazmatskog retikuluma. U tim su slučajevima vakule obučene proteinima policajca I-kompleksa. Vjeruje se da se na taj način vraćaju različiti federalni enzimi i receptori u membranama. Ove karakteristike ponašanja transportnih vezika dale su osnovu hipoteze o postojanju dvije vrste transportnih komponenti AG-a. Prema jednom od njih, najrasniji, u AG-u postoje stabilne membranske komponente, na koje su tvari uz pomoć transportnih vakuela prenose iz estefa. Prema alternativnom modelu AG je dinamičan derivat ER: membranskih vakula sa prijateljem sa novim CISTErom, koji se zatim kreću kroz cijelu zonu i na kraju prelazi u transportne vezikule. Prema ovom modelu, retrogradni policajac i veSexu vraćaju trajne proteine \u200b\u200bAG u mlađe tenkove. Stoga se pretpostavlja da je tranzicijska zona ER "porodiljska bolnica" za Golgi aparat. |
Pitanje 37. Lizosoma. Funkcija obrazovne strukture. Heterogenost lizosomi. Patologija Lysosomes.
Lizosoma - Ćelijski organoloid dimenzija 0,2 - 0,4 μm, jedna od vrsta vezikula. Ovi jednobojni organsiri dio su vakuuma (endometrijski sustav ćelija). Različite vrste Lizosomi se mogu smatrati zasebnim staničnim pretincima.
Funkcije Lysosomes su:
· Probavka zarobljene ćelije tokom endocitoze tvari ili čestica (bakterije, druge ćelije)
· Autofagija - uništavanje nepotrebnih ćelija struktura, na primer, tokom zamene starih organoida nove, ili probave proteina i drugih tvari proizvedenih unutar same ćelije
· Avtoliz - Samosejanje ćelije koja vodi do njegove smrti (ponekad ovaj proces nije patološki i prati razvoj tijela ili diferencijacije nekih specijalizovanih ćelija). Primjer: Kada pretvorite na glavu u žabu, lizosomi koji su u uskim ćelijama, probaviti: rep nestaje, a tvari formirane tokom ovog procesa apsorbiraju i koriste druge stanice.
Ponekad zbog netačan rad Lizosoma razvija akumulacijske bolesti u kojima enzimi zbog mutacija ne rade ili ne rade loše. Primjer akumulacijskih bolesti može poslužiti kao amavrotički idioti prilikom akumuliranja glikogena.
· Jaz lizosoma i izlaz na hiloplazmu podijeljenih enzima prati je oštrim povećanjem njihove aktivnosti. Takvo povećanje aktivnosti enzima primijeće se, na primjer, u žarištu nekroze sa infarktom miokarda i pod djelovanjem zračenja.
Lizosomi se formiraju od mjehurića (vezika) odvojenih od Golgi aparata i mjehurića (endos), u kojim supstancima padaju tokom endocitoze. U formiranju autolizma (autofagos), membrane endoplazmatskih retikuluma sudjeluju. Svi proteini lizosoma sintetiraju se na "sjedećim" ribosomima na vanjskoj strani membrane endoplazmatskog retikuluma, a zatim prolaze kroz svoju šupljinu i kroz Golgi aparat.
Lizosomi su heterogene organele s različitim oblicima, dimenzijama, ultrastrukturnim i citohemijskim karakteristikama. "Tipični" lizosi životinjskih ćelija obično imaju dimenziju od 0,1-1 μm, sferični ili ovalni oblik. Broj lizosoma varira od jedne (velike vakuole u mnogim biljkama i stanicama gljiva) do nekoliko stotina ili hiljada (u životinjskim ćelijama).
Ne postoji općenito prihvaćena klasifikacija i nomenklatura za različite faze zrenja i vrsta lizosoma. Razlikovati primarne i sekundarne lizosome. Prvi se formiraju u području Golgi uređaja, oni imaju enzime u neaktivnom stanju, drugi sadrži aktivne enzime. Obično se enzimi lizosoma aktiviraju spuštanjem pH. Među Lysosomima se mogu izolirati i heterolisosomi (probaviti materijal koji ulazi u ćeliju vani - phaguo- ili pinocitozom) i autolisosomima (uništavajući vlastite proteine \u200b\u200bili ćelije). Najčešće korištena je sljedeća klasifikacija lizosoma i srodnih odjeljaka:
- Rano endosome - endocitoza (pincitomic) mjehurići ulazi u to. Od ranog endosoma, receptori koji su im dali njihov teret (zbog smanjenog pH) vraćaju se u vanjsku membranu.
- Kasni endosome - u svojim ranim endosomima postoje mjehurići sa materijalom apsorbiran u pinocitozi i mjehurići iz Golgi aparata sa hidrolilama. Manneco-6-fosfatni receptori vraćaju se sa kasnih endosoma na Golgi aparat.
- Lysosome - u njemu iz kasnih endosoma postoje mjehurići sa mješavinom hidroliza i probavljivog materijala.
- Fagomija - veće čestice (bakterije itd.) Ulaze u njega, apsorbirane fagocitozom. Fagosome su obično spojeni sa lizosom.
- Autofagosome - okružen dve membrane, delom citoplazme, obično uključujući bilo koji organsa i formiran tokom makrostofagije. Spaja se sa lizosom.
- Multisikularna telad obično su okružena jednom membranom, sadrže unutarnje okružene mjerževe membrane. Formirani su kao rezultat procesa koji nalikuje mikroutofa (vidi dolje), ali sadrži materijal dobiven izvana. U malim mjehurićima, vanjski membranski receptori obično su izloženi, a zatim (na primjer, receptori faktora epidermalnog rasta). U fazi formiranja odgovara ranom endosomi. Opisano je formiranje više-mezzutarnih Bika, okruženo dvije membrane, pupljenjem iz nuklearne ljuske.
- Preostali Bik (TELICO) - Mjehurići koji sadrže neiskoređeni materijal (posebno lipofuscin). U normalnim ćelijama se spajaju s vanjskim membranom i egzocitozom napuštaju ćeliju. U starenju ili patologiji akumuliraju.
Pitanje 38. Opišite put iz sekretarnog proteina sa stranice sinteze proteina prije izlaska na ćeliju.
U ćelijama u kojima se sekrecija javlja kao odgovor na vanćelijski signal, izlučene proteine \u200b\u200bsu koncentrirani i pohranjeni u sekretarnim mjehurićima (često se nazivaju tajničkim granulama zbog mračne jezgre). Prilikom dobijanja odgovarajućeg signala oslobađaju ih expocitozom. Izvanredni mjehurići su izvedeni iz mreže Golgi Trans. Vjeruje se da se za njihovo formiranje, klantinke i pridružene proteine \u200b\u200bstvaraju "mjehurić", jer je dio površine formiranja mjehurića obično prekriven Clawinom. Ova granica uklanja se ubrzo nakon što se mjehurić u potpunosti formira (Sl.8-76).
Poput lizosoma hidrolaza, proteini namijenjeni sekretarnim mjehurićima (oni se često nazivaju sekretarnim proteinima) moraju se odabrati i pakirati u odgovarajuće mjehuriće u GOLGI TRANS mreži. Očigledno, u ovom slučaju se događa selektivna agregacija sekretornih proteina. Rezultirajući agregati u elektrokonomskom mikroskopu izgledaju kao materijal za elektronsko moždani udar u GOLGI TRANS mreži. "Signal za sortiranje", vodeći proteine \u200b\u200bu takve agregate, nije poznat, ali očigledno je ovo signalna parcela zajednička za mnoge sekretarne proteine. Takav zaključak potvrđuje se sljedećim podacima: ako se gen kodiraju iz sekretarnog proteina prebačen u sekretnu ćeliju drugog tipa, u normalnom proteinu, ne sintetizirajući ovaj protein, tada će se tuđi protein biti pakiran i u sekretarnim mjehurićima.
Nije poznato kako, u formiranju sekretornih mjehurića, biraju se agregati koji sadrže sekretarnje proteine. Sekretorirani mjehurići imaju jedinstvene membranske proteine, od kojih neki mogu poslužiti kao receptori (u prekomjernoj mreži Golgi) da bi se obvezao agregirani materijal za pakiranje. Sekretori mjehurići su više mjehurića koji nose lizosoma i jedinice koje su sadržane u njima su prevelike tako da se svaka molekulacija izlučenog proteina može obratiti receptoru u mjehurićima mjehurića, jer se to događa tokom transporta lizosoma enzima. Snimanje ovih agregata sekretornih granula prilično podsjeća na apsorpciju čestica sa fagocitozom na staničnoj površini, koja se događa i sa sudjelovanjem obloženih membrana.
Nakon nezrelih sekretarskih mjehurića iz GOLGI TRANS MREŽE, oni gube kaima, a njihov je sadržaj snažno koncentriran. Takva kondenzacija javlja se oštro i može biti uzrokovana zakiseljujućim srednjom u šupljini mjehurića zbog rada PUMP-a ovisne o ATP-u u svojoj membrani. Agregiranje tajnih proteina (ili drugih komponenti) i njihova naknadna kondenzacija u sekretornim mjehurićima uzrokuje porast koncentracije tih proteina u 200 puta u odnosu na Golgi aparat. Zahvaljujući tome, sekretarnji mjehurići imaju priliku osloboditi velike količine materijala u timu.
Pitanje №39.opy Put hidroliza sa mjesta njihove sinteze do odredišta.
Hidrolaza, Klasa enzima katalizacije hidrolize. Oni mogu djelovati na estera i glikozidnim vezama, u vezi s jednostavnim eterom. S-s u sulfidima, C-n u kućnom ljubimcu, i drugi.
HidrolazaHidroliza esterskih obveznica (esterase) aktivira se na esterima karboksilnih i thio-karboksilnih kiselina, monoestera fosforne kiseline itd. Za ovu potklasu uključuju, posebno enzimi koji igraju važnu ulogu u metabolipidima. Nukleinske kiseline i nukleozidi. Na primjer, arilsulfataza , acetilcholinesteraza , deoxyribonucleasease . lipase , fosfataza , Phosfoendodoxocyribonuclease lipaze
Enzimi kataliziranje hidrolize c-N veze U peptidima i proteinima (peptidhidrolaze), najbrojnija grupa hidrolaza Oni uključuju enzime koji ukidaju jednu ili dvije aminokiseline sa N- ili C-Terminusom polipeptidnog lanca (na primjer, aminoptidaze , karboksipeptidaza ), kao i endopeptidaze ili proteinaze, dijeljenje lanca daleko od krajnjeg salda. Peptidhidrolaze igraju važnu ulogu ne samo u katabolizmu proteina i peptida, već i u biolu. Uredba (hormonalna regulacija, aktiviranje pro-procjene, regulacija krvnog tlaka i metabolizma soli itd.).
Pitanje 40. Opišite put makromolekule od trenutka primitka u ćeliju do asimilacije.
znati
Pitanje 41. Uloga AG-a i ER u regeneraciji i ažuriranju površinskog aparata ćelije (PAC)
Uloga AG-a u ažuriranju KAP-a:
Golgi aparat. U mnogim životinjskim ćelijama, na primjer, nervozno, ima oblik složene mreže koja se nalazi oko kernela. U ćelijama Rayanijuma i najjednostavnije, Golgi aparat predstavljen je zasebnim venama srpa ili valjka. Struktura ovog organa slična je ćelijama biljnih i životinjskih organizma, uprkos raznolikosti njegovih oblika.
Golgi aparat uključuje: šupljine ograničene na membrane i grupe od 5-10); Veliki i fini mjehurići smješteni na krajevima šupljina. Svi ovi elementi čine jedan kompleks.
Golgi aparat obavlja mnogo važnih funkcija. Na kanalima endo-plazme mreže, proizvodi se od sintetičke aktivnosti ćelija - proteini, ugljikohidrati i masti prevože se na njega. Sve ove supstance spavanja - a zatim se nakupljaju, a zatim u obliku velikih i malih mjehurića ulaze u citoplazmu ili se koriste u samoj ćeliji tijekom sredstava za život ili su izveli iz njega i koriste se iz njega i koriste se u tijelu. Na primjer, u ćelijama gušterače sisara sintetiziraju se enzimi za probave, koji se nakupljaju u šupljinama organa. Tada se formiraju mjehurići ispunjeni enzimima. Izvedeni su iz ćelija u kanalima od gušteracije, odakle teku u šupljinu creva. Druga važna funkcija ovog organa je da sinteza masti i ugljikohidrata (polisaharidi), koji se koriste u ćeliji i koji su uključeni u membranu, javljaju se na njegovim membranama. Zahvaljujući aktivnostima Golgi aparata, ažuriranja i rasta pasulja u plazmi.
(Pogledajte dolje i o AG i o ER, 2 drugih izvora).
Uloga ER-a u ažuriranju PAK:
Endoplazmatski reticulum (Endoplasminatska mreža) je sistem rezervoara, cevastih i vakula, ograničenog citomembrana. Razlikovati granularno (grungy) i agranularno (glatko) endoplazmatsko retikulum; Ravne gomile prevladavaju u prvim - tenkovima, u drugom - tubule. Membrane grube retikuluma na strani hiloplazme prekrivene su ribosomima. Stupanj razvoja ovog organaloga ovisi o nivou metaboličke aktivnosti i diferencijacije K.: Jači je od ćelija u ćelijama, aktivno sintetiziraju proteine.
(Još jedan izvor).
ER - Transportni proteini.
Er šupljina odvojena je od citoze jedne membrane ( Membrana ), Veza za obvezuju između dva odjeljka. Naprotiv, er šupljine i svaki rezervoar Golgi odvojeni su jedni od drugih po dvije membrane i citosole, pa se prevoz makromolekula između tih organele vrši pomoću transportnih mjehurića.
Svi novoisetizirani proteini, bez obzira na njihovo odredište (er šupljina, golgi, lizozoma ili vanćelijski prostor), prvo dođu u er šupljinu.
Neki proteini se kreću od citosola u Grungy ER odmah nakon njihove sinteze.
Ovo su proteini dvije vrste:
1) Tječnice koji su samo djelomično prenose kroz er membranu i ostaju zatvoreni u njemu i
2) topiv u vodi, koji se u potpunosti prenosi kroz er membranu i prekriveni su u svojoj šupljini.
U ćelijama sisavaca počinje uvoz proteina u ER-u prije nego što se lanac polipeptida u potpunosti sintetizira, I.E. javlja se istovremeno s emitovanjem (cotranslova).
Dakle, u citoplazmi postoji dva prostorno izolirana populacija ribosoma. Neki od njih (ribosomi povezani sa membranom) nalaze se na er membranskoj površini okrenutoj citoplazmi i bave se sintezom proteina, koji se odmah prebacuju u ER. Ostalo (ribosomi su besplatni) ne odvojene u bilo koju membranu i proizvode sve ostale proteine \u200b\u200bkodirane od jezgre. Srodni i slobodni ribosomi su identični u strukturi i funkciji. Oni se razlikuju samo u proteinima koji su u svakom trenutku sintetizirani na njima. Ako ribosom dobije sintezu proteina sa signalnim peptidom za ER, tada takav signal šalje ribosome u er membranu.
(Još jedan izvor).
Već smo naglasili koliko su opsežne strukture endoplazmatskog retikuluma i golgi aparat U sekretarnim ćelijama. Osnova ovih struktura su membrane iz lipida Bislowsa, slične strukturi iz membrane ćelije. Zidovi membrana sadrže enzime koji kataliziraju sintezu mnogih tvari koje zahtijeva ćeliju.
Većina sintetičkih procesa dolazi u endoplazmatskom retikulumu. Ovdje formirane tvari šalju se u Golgi aparat, gdje su podvrgnute daljnjoj obradi prije nego što uđu u citoplazmu. U početku se treba zaustaviti na tvarima koje su sintetizirane u zasebnim područjima retikuluma i Golgi uređaja.
Sinteza proteina na grubi endoplazmatski reticulum. Na vanjskoj površini grubog endoplazmatskog retikuluma sadrži veliki broj pričvršćenih ribosoma na njemu; Na njima se pojavljuje sinteza proteina, od kojih je mali iznos u citosolu, a glavni dio je u lumen cijevi i mjehurićima Reticulum, tj. u endoplazmatskoj matrici.
Sinteza lipida u glatkom endoplazmatskom retikulumu. Endoplazmatski reticulum može sinteza lipida, posebno fosfolipida i holesterola. Oni se brzo rastvaraju u membranom bisalu, što doprinosi daljnjem rastu struktura retikuluma, uglavnom glatkih.
Mali mjehurići, nazvan prijevoz, ili ER-vakulacija se neprestano odvajaju od glatke retikulum membrane, čime se na taj način sprečava prekomjerni rast. Većina ovih vakula za vozila se zatim brzo kreće u Golgi aparat.
Ostale funkcije funoplazmatske retikulum. Endoplazmatski retikulum, posebno gladak, ima i druge važne funkcije.
1. Omogućavanje enzima koji dijele glikogen ako je potrebna energija.
2. Omogućavanje velikog broja enzima koji mogu oštetiti tvari štetne za ćelije, na primjer, lijekovi. Metode neutralizacije uključuju koagulaciju, oksidaciju, hidrolizu, spoj sa glukuronskim kiselinom itd.
Struktura kompleksa Golgi
Golgi kompleks (Kg), ili interni neto aparat - Ovo je poseban dio metaboličkog sistema citoplazme, sudjelovanje u procesu izolacije i formiranja konstrukcija membranske ćelije.
CG se može videti u optičkom mikroskopu kao rešetka ili zakrivljenih teladi u obliku štapa leže oko kernela.
Pod elektronskim mikroskopom otkriveno je da je ova organa predstavljena tri vrste formacija:
Sve komponente Golgi uređaja formiraju glatke membrane.
Napomena 1.
Povremeno ag ima strukturu metre i nalazi se u blizini jezgre u obliku poklopca.
AG se nalazi u svim ćelijama biljaka i životinja.
Napomena 2.
Golgi aparat značajno se razvija u sekretornim ćelijama. Posebno je dobro vidljiv u nervnim ćelijama.
Unutarnji intermogramski prostor ispunjen je matricom, koji sadrži određene enzime.
Golgi aparat ima dvije zone:
- zona formiranjagdje uz pomoć Vesikula dolazi s materijalom koji se sintetizira u endoplazmatskoj mreži;
- zona sazrijevanjagdje se formiraju tajne i sekretarne torbe. Ova tajna se nakuplja na terminalnim dijelovima AG-a, odakle su sekretarnjostekni izleti. U pravilu, takvi veslisti nose tajnu izvan ćelije.
Lokalizacija kg.
U apolarnim ćelijama (na primjer, nervozno) kg se nalazi oko kernela, u sekretoru se odvija između jezgre i apikalnog pola.
Kompleks golgih torbi ima dvije površine:
formativan (nezreli ili regenerator) CIS-površina (od lat. CIS - s ove strane); funkcionalan (zrelo) - trans površina (od lat. Trans - za) za).
Golgiov stupac sa svojom konveksnom formativnom površinom okrenutom jezgri, uz zrno endoplazmatsku mrežu i sadrži male okrugle mjehuriće zvani posrednik. Zrela površina konkavne vrećice okreće se prema vrhu (apikalni stup) ćelije i završava velikim mjehurićima.
Formiranje GOLGI kompleksa
Membrane kg sintetiraju se sa zrnatom endoplazmatskim mrežom koja je u blizini kompleksa. Susjedni dijelovi EPS-a gube ribosomi, mali, takozvani, takozvani transport ili srednje vezike. Prelaze na formativnu površinu Golgi panjeva i spajaju se svojom prvom torbom. Na suprotnoj (zreli) površini kompleksa Golgji nalazi se vreća pogrešnog oblika. Njegova ekspanzija je objektivne granule (kondenzacijska vakula) - kontinuirano računati i pretvoriti u mjehuriće ispunjene tajnom - sekretarnim granulama. Dakle, u mjeri membrana zrele površine kompleksa na sekretornim vezici, gomile formativne površine nadopunjuju se zbog endoplazmatske rešetke.
Funkcije kompleksa Golgi
Glavna funkcija Golgi uređaja je uklanjanje sintetiziranih ćelija tvari. Te su tvari prevožene kroz ćelije endoplazmatske mreže i akumuliraju se u mjehurićima mrežnog aparata. Tada su ili su zacrtani vanjsko okruženje Ili ih ćelija koristi u procesu života.
Kompleks također koncentrira neke tvari (na primjer, boje), koje dolaze u ćeliju vani i moraju se izvući iz nje.
U biljnim ćelijama, kompleks sadrži enzime sinteze polisaharida i samog polisaharidnog materijala koji se koristi za izgradnju granata kavezu celuloze.
Pored toga, kg sintetizira te hemikalije koje tvore ćelijsku membranu.
Općenito, aparat Golgi izvodi takve funkcije:
- akumulacija i izmjena makromolekula, koji su sintetizirani u endoplazmatskoj mreži;
- formiranje složenih tajni i sekretornih vezikula kondenzacijom sekretornog proizvoda;
- sinteza i modifikacija ugljikohidrata i glikoproteina (glikočicalizi, sluz);
- izmjena proteina - dodavanje polipeptid raznih kemijskih formacija (fosfat - fosforil, karboksilacija - karboksilacija), formiranje složenih proteina (lipoproteini, glikoproteini, sluznice) i cijepanje polipeptida;
- važno je za formiranje, ažuriranje citoplazmatske membrane i ostale membranske formacije zbog formiranja membranskih oprega, koji se dodatno spojiju sa ćelijom membrana;
- formiranje lizosoma i specifične zrnate u leukocitima;
- formiranje peroksisa.

Protein i dijelom, Carbohidrat Sadržaj KG dolazi s zrnato endoplazmatskim mrežicom, gdje se sintetizira. Glavni dio komponente ugljikohidrata formiran je u vrećama kompleksa s sudjelovanjem glikoziltransferaza enzima, koji su u membranama vreća.
U kompleksu gomila ćelija se sadrže tajne koje sadrže glikoproteides i glikozaminoglycans konačno su formulisane. U kg se izmirnici zrele, koje se uvlače u mjehuriće, a kretanje ovih mjehurića u smjeru plazmalemme Konačna faza izlučivanja je guranje formirane (zrele) vestitule izvan ćelije. Uklanjanje sekretornih inkluzija iz ćelije vrši se namotavanjem mjehurnog mjehurića u plasmalamm i izbor sekretornih proizvoda izvan ćelije. U procesu pokretnih sekretarskih mjehurića na apikalni pol membranskih ćelija, zgušnjavaju se od početnog 5-7 Nm, dostižući debljinu plasmalamina 7-10 Nm.
Napomena 4.
Postoji međuovisnost između ćelijskog aktivnosti i dimenzija Golgji kompleksa - sekretne ćelije imaju velike stupce kg, dok ne-rudar sadrži mali broj torbi kompleksa.
Organoid. - Trajno, nužno prisutno, ćelijske komponente koje vrše određene funkcije.
Endoplazmatski reticulum
Endoplazmska mreža (EPS), ili endoplazmatski reticulum (EPR)- Jednotamni organoid. To je sustav membrana koji formiraju "tenkove" i kanale povezani jedni s drugima i ograničavajući jedan unutarnji prostor - EPS šupljina. Membrane s jedne strane povezane su s citoplazmatskim membranom, s druge, sa vanjskim nuklearnim membranom. Postoje dvije vrste EPS: 1) grubo (zrnalo), koje sadrže ribosome na njenoj površini i 2) glatka (agranularna), čija membrane ne prevoze ribosomima.
Funkcije: 1) Vozila supstanci iz jednog komada ćelija u drugu, 2) odvajanje citoplazmatskih ćelija za odjeljke ("odjeljke"), 3) sinteza ugljikohidrata i lipida (glatki EPS), 5) sinteza proteina (grubi EPS), 5) Mjesto formiranja Golgi uređaja.
Ili golgi kompleks- Jednotamni organoid. To je snop spljoštenih "tenkova" sa proširenim ivicama. S njima je povezan sistem malih montiranih mjehurića (Golgi Bubbles). Svaka bička obično se sastoji od 4-x-6 "tenkova", strukturalna je funkcionalna jedinica Golgi uređaja i naziva se doktoma. Dondyom broj u ćeliji se kreće od jednog do nekoliko stotina. U biljnim ćelijama, razdvajanje su odvojeni.
Uređaji za golgi obično se nalazi u blizini ćelijskog jezgra (u životinjskim ćelijama često u blizini mobilnog centra).
Funkcije Golgi uređaja: 1) Akumulacija proteina, lipida, ugljikohidrata, 2) Primljena organskih tvari, 3) "ambalaža" u membranskim mjehurićima proteina, lipida, ugljikohidrata, 4) izlučivanje proteina, lipida, ugljikohidrata, 5) sinteza ugljikohidrata i Lipidi, 6) Lysosomes. Sekretna funkcija je najvažnija, tako da je Golgi aparat dobro razvijen u sekretornim ćelijama.
Lysosomes
Lysosomes - Organoidi sa jednim gramama. Predstavite male mjehuriće (promjer od 0,2 do 0,8 mikrona) koji sadrži skup hidrolitičkih enzima. Enzimi su sintetizirani na grubim EPS-om, premještene u Golgi aparat, gdje se pojavljuju njihova izmjena i ambalaža u membranskim mjehurićima, koji nakon odvajanja od Golgi aparata postaju zapravo lizosomi. Lysosome može sadržavati od 20 do 60 različitih vrsta hidrolitičkih enzima. Cijepanje tvari uz pomoć enzima poziva lis.
Razlikovati: 1) primarni lizosomi, 2) sekundarni Lysosomes. Primarni se nazivaju lizosomi, odlaze se iz Golgi aparata. Primarni lizosomi su faktor koji pruža egzocitozu enzima iz ćelije.
Sekundarno se nazivaju lizosomi formiranim fuzijama primarnih lizosoma sa endocitozom vakuole. U ovom slučaju postoje probave tvari koje su u ćeliju ušle po fagocitozi ili pinocitozi, pa se mogu nazvati probavnim vakulama.
Autofagi. - Proces uništavanja nepotrebnih ćelija struktura. Prvo, struktura koja treba uništiti je okružena jednom membranom, tada se rezultirajuća membranska kapsula spaja sa primarnim lizosom, formiran je i sekundarni lizing (autofagični vakuol) u kojem se ta struktura probavlja. Proizvodi za probavu apsorbiraju citoplazme, ali dio materijala ostaje netaknut. Sekundarni Lysosome koji sadrži ovaj neizostavljeni materijal naziva se preostalo tijelo. Exocitozom, neovlaštene čestice se uklanjaju iz ćelije.
Autoliz - Samouništavanje ćelije koja se događa zbog oslobađanja sadržaja lizosoma. Obično se autolizacija odvija u metamorfozi (nestanka repa od glave žaba), involucija maternice nakon porođaja, u žarištu tkiva.
Funkcije Lysosomes: 1) unutarćelijska probava organskih tvari, 2) uništavanje nepotrebnih mobilnih i ne-mobilnih struktura, 3) sudjelovanje u procesima reorganizacije ćelija.
Vakuola
Vakuola - Organizova s \u200b\u200bjednim gramom su "kapaciteta" ispunjeni vodena rješenja Organske i anorganske tvari. EPS i Golgi aparat sudjeluju u formiranju vakula. Mlade biljne ćelije sadrže mnogo malih vakuola koje su tada, kao diferencijacije i diferencijacije ćelija, stanice se međusobno spajaju i oblikuju jednu veliku centralni vakolol.. Centralni vakuola može trajati do 95% količine zrele ćelije, kernel i organoidi se guraju u staničnu školjku. Membrana ograničavanje vakula povrća naziva se tonoplast. Tečno punjenje vegetativnog vakuola, pozvan sok za ćelije. Cellular sok uključuje organske i neorganske soli topljivog i neorganskih soli, monosaharida, disačarida, aminokiseline, konačne ili toksične metaboličke proizvode (glikozide, alkaloide), neke pigmente (antocijani).
U životinjskim ćelijama su male probavne i autofagične vakule vezane za grupu sekundarnih lizosoma i koji sadrže hidrolitičke enzime. Jednoćelijske životinje imaju više kontraktilnih vakuela koje obavljaju funkciju akrivezacije i odabira.
Značajke vakuole: 1) Akumulacija i skladištenje vode, 2) regulacija metabolizma vodenog soli, 3) održavanje tlaka iz turneje, 4) akumulacija metabolite u vodi, rezervnih hranjivih sastojaka, 5) bojanje boja i voća i privlačenje oprašivača i distributera Sjemenke, 6) cm. Funkcije Lysosomes.
Endoplazminatska mreža, golgi, lizosomi i vakulijski obrazac jedinstvena vakularna ćelijska mreža, čiji su neki elementi od kojih mogu ići jedni drugima.
Mitohondria

1 - vanjska membrana;
2 - unutrašnja membrana; 3 - matrica; 4 - Crista; 5 je multimenski sistem; 6 - Prsten DNK.
Oblik, dimenzije i broj mitohondrije se razlikuju izuzetno. U obliku mitohondrije može se ukinuti, zaobljena, spirala, cupid, razgranati se. Dužina mitohondria kreće se od 1,5 do 10 mikrona, promjera - od 0,25 do 1,00 mikrona. Količina mitohondrije u ćeliji može dostići nekoliko hiljada i ovisi o metaboličkoj aktivnosti ćelije.
Mitohondria je ograničena na dvije membrane. Vanjska membrana mitohondrija (1) glatka, unutarnja (2) formira brojne nabore - crysto (četiri). Crystys povećavaju površinu unutarnje membrane, na kojoj se nalaze multimenzijski sustavi (5) koji se nalaze u sintezi ATP molekula. Unutarnji prostor mitohondrije ispunjen je matricom (3). Matrica sadrži prsten DNK (6), specifičan IRNK, ribosomi prokariotskog tipa (70-ih), Krebs ciklus enzima.
Mitohondrial DNK nije povezan sa proteinima ("golim"), pričvršćenim na unutrašnju membranu mitohondria i nosi informacije o strukturi od oko 30 proteina. Za izgradnju mitohondrije potreban je mnogo više proteina, tako da su informacije o većini mitohondrijskih proteina sadržane u nuklearnoj DNK, a ovi proteini sintetizirani su u citoplazmi u citoplazmi. Mitohondria je sposobna da se autonomno pomnoži podjelim u dva. Između vanjskih i unutrašnjih membrana nalazi se proton rezervoargdje se događa akumulacija H +.
Funkcije Mitohondria: 1) sinteza ATP, 2) Cjepavanje kisika organskih tvari.
Prema jednoj od hipoteze (teorija simbiogeneze), mitohondria se dogodila od drevnih slobodnih živih aerobičnih prokarnotskih organizama, koji slučajno prodire u hostu. Sledeći podaci dokazuju se u korist ove hipoteze. Prvo, mitohondrijski DNK ima iste karakteristike strukture, kao i DNK modernih bakterija (zatvoreno u prstenu, a ne povezane sa proteinima). Drugo, mitohondrijski ribosomi i ribosomi bakterija pripadaju jednoj vrsti - 70-ih. Treće, mitohondrijski mehanizam podjele sličan je tim bakterijama. Četvrto, sinteza mitohondrijskih i bakterijskih proteina potiskuje isti antibiotici.
Plastici

1 - vanjska membrana; 2 - unutrašnja membrana; 3 - Strom; 4 - tilakoidni; 5 - grana; 6 - Lammella; 7 - zrna škroba; 8 - LIPID kapi.
Zemljišta karakteristična su samo za biljne ćelije. Razlikovati tri glavne vrste plastike: leukoplasts - bezbojni plastidi u ćelijama neobožavanih dijelova biljaka, kromoplasti - obojeni plasti obično žute, crvene i narančaste boje, hloroplasti - zeleni plasti.
Kloroplasti. U ćelijama vilje biljke Hloroplasti imaju oblik leća poput bikona. Dužina hloroplasta kreće se od 5 do 10 mikrona, promjera - od 2 do 4 mikrona. Hloroplasti su ograničeni na dvije membrane. Vanjska membrana (1) je glatka, unutarnja (2) ima složenu sklopljenu konstrukciju. Naziva se najmanji preklop tilakoid (četiri). Telakoidna grupa postavljena kao hrpa kovanica se zove grana (pet). U kloroplastu sadrži prosječno 40-60 grafilova. Grairi su povezani međusobno sa sakupljenim kanalima - lammella (6). Tylacoid membrane su ugrađeni fotosintetički pigmenti i enzimi koji pružaju sintezu ATP-a. Glavni fotosintetski pigment je hlorofil, koji određuje zelenu boju kloroplasta.
Ispunjen je unutarnji prostor kloroplasta stroma (3). U stromi su prsteni "goli" DNK, 70-ih Ribosomi tipa, kalvine ciklus enzima, zrno škroba (7). Unutar svakog tilakoida dolazi u rezervoar protona, H + akumulacija. Hloroplasti, kao i mitohondria, sposobni su za autonomnu reprodukciju dijeljenjem u dva. Nalaze se u ćelijama zelenih dijelova viših postrojenja, posebno mnogo hloroplasta u lišću i zelenim plodovima. Hloroplasti nižih biljaka nazivaju se kromatofora.
Funkcija kloroplasta: fotosinteza. Vjeruje se da su se kloroplasti dogodio od drevnih endosimbiotskih cijanobakterija (teorija simbogeneza). Osnova za takvu pretpostavku je sličnost kloroplasta i modernih bakterija za brojne znakove (bučne, "gole" DNK, 70-ih Ribosomi tipa, metoda reprodukcije).
Leukoplasts. Obrazac varira varira (sfernim, zaobljenim, šoljicama itd.). Leukoplasti su ograničeni na dvije membrane. Vanjska membrana je glatka, unutrašnji formira male tilakoide. U stromi su prsteni "goli" DNK, ribosomi tipa 70-ih, enzimi sinteze i hidroliza rezervnih hranjivih sastojaka. Pigmenti su odsutni. Posebno mnogi leukoplasti imaju ćelije podzemnih organa postrojenja (korijenje, gomolji, rizomi itd.). Funkcija leukoplasta: Sinteza, akumulacija i skladištenje rezervnih hranjivih sastojaka. Amyloplasts - leukoplasti koji sintetišu i akumuliraju škrob, elayoplasts - ulja, proteinoplasti - Proteini. U istom leukoplastu može akumulirati različite tvari.
Hromoplasti. Ograničene sa dvije membrane. Vanjska membrana je glatka, unutrašnja ili također glatka ili formira pojedinačne tilakoide. U stromi se nalaze prstene i pigmenti - karotenoidi koji daju kromoplastike žute, crvene ili narančaste boje. Oblik akumulacije pigmenata je različit: u obliku kristala, rastvorenih u lipidnim kapima (8), a drugi nalaze se u ćelijama zrelog voća, latica, jesenjih listova, rijetko korijen. Hromoplasti se smatraju završnom fazom razvoja plastike.
Funkcija kromoplasti: Bojanje boja i voća i na taj način privlačeći oprašivače i distributera sjemena.
Sve vrste plastida mogu se formirati iz precipitala. Proklizaci - Mali organoidi sadržani u meristematičnim tkivima. Budući da plastidi imaju opće porijeklo, među njima su moguća za međusobno povezivanje. Leukoplasti se mogu pretvoriti u kloroplaste (zelenilo gomolji krumpira u svjetlu), hloroplasti - u kromoplastima (žutilo od lišća i crvenilo voća). Pretvaranje kromoplasta u leukoplastima ili hloroplastima smatra se nemogućim.
Ribosomi

1 - velika podjedinica; 2 - Mala podjedinica.
Ribosomi - Nemikrijumčareni organi, prečnik od oko 20 Nm. Ribosomi se sastoje od dvije podjedinice - velike i male, koje se mogu disocirati. Hemijski sastav Ribosomi - proteini i rrna. Molekule RRNA su 50-63% mase ribosoma i formiraju svoj strukturni okvir. Dvije vrste ribosoma su ukinute: 1) eukariotic (sa cjelokupnom ribosomim konstante sedimentacije - 80-ih, malu podjedinicu - 40-ih, velikih - 60-ih) i 2) prokariotski (70-ih, 30-ih, 50-ih godina.
Kao dio ribosome eukariotskog tipa 4 RRNA molekula i oko 100 proteinskih molekula, prokariotskog tipa - 3 molekule RRNA i oko 55 molekula za oko 55 proteina. Tokom biosinteze, ribosoma protein može "raditi" po jednu ili ujediniti u komplekse - poliribosomi (Polisomas). U takvim kompleksima povezani su jedni s drugima istim molekulom s tintom. Prokarnotičke ćelije imaju samo 70-ih ribosoma tipa. Eukariotske ćelije imaju ribosomi poput 80-ih (grouse membrane EPS, citoplazma) i tipa 70-ih (mitohondria, hloroplasti).
Eukariot Ribosome podjedinice formiraju se u nukleolinu. Kombinovanje podjedinica u cjelinu ribosoma javlja se u citoplazmi, u pravilu, tokom biosinteze proteina.
Funkcija ribosoma: Montaža lanca polipeptida (sinteza proteina).
Cytoskeletton
Cytoskeletton Obrazovani mikrotubima i mikrofilamentima. MicroTubule - cilindrične neubrancije. Dužina mikrokula kreće se od 100 μm do 1 mm, promjer je otprilike 24 nm, debljina zida je 5 nm. Glavna hemijska komponenta je tubulin protein. Mikrotubule su uništeni pod utjecajem kolkhicina. Mikrofilamenti - niti s promjerom od 5-7 Nm, sastoje se od djela proteina. Mikrotubule i mikrofilamenti formiraju složene preplitanje u citoplazmi. Funkcije citoskeleta: 1) Definicija oblika ćelije, 2) podrške organoidima, 3) formiranje vretena divizije, 4) učešće u pokretima ćelije, 5) trenutnu organizaciju citoplazma.

Sadrži dva centriole i Centrofer. Centril To je cilindar, čiji zid formiraju devet grupa od tri prosuta mikrotubula (9 trojke) međusobno povezane kroz određene intervale križnih shyshiva. Centriole se kombiniraju u par gdje se nalaze pod pravim uglom jedni drugima. Prije razdvajanja ćelija, centrirani su se odvijali suprotnim polovima, a podružnica centra nastaje u blizini svake od njih. Oni čine dindleske podjele koje doprinose ujednačenoj distribuciji genetski materijal između ćelija kćeri. U ćelijama viših biljaka (doseljenih, obloženih slanicom), ćelijski centar centrioleja nema. Centrioli se odnosi na samoproduktiranje citoplazme, oni nastaju kao rezultat dupliranja već dostupnih sredi. Funkcije: 1) Osiguravanje odstupanja hromozoma na ćelijske stupove tokom mitoze ili mejoze, 2) središte organizovanja citoskeleta.
Motion Organis
Prisutni ne u svim ćelijama. Oprema pokreta uključuje Ciliju (Infuzorija, respiratorni epitel), flagela (FAZELON, SPEMATOZOA), FALSE I TABELI (NUTS, LEUKOCYTES), Myofibrils (mišićne ćelije), itd.
FEDELA I CILIA - Organoidi filamentalnog oblika su asonsee, ograničena membrana. Axona - cilindrična struktura; Zid cilindra formira se za devet pari mikrotubula, u svom centru postoje dva pojedinačna mikrotubula. U podnožju osi postoje bazalne telad, zastupljene su dva uzajamno okomita centra (svaki bazalni pozivatelj sastoji se od devet trojki mikrotubula, u svom centru nema mikrotubula). Duljina boka doseže 150 μm, Cilia je nekoliko puta kraće.
Miofibrils Sastoje se od actin i samih miofilamenata koji smanjuju mišićne ćelije.
Idi predavanja broj 6. "Eukariotska ćelija: citoplazma, ćelijska školjka, struktura i funkcija ćelijskih membrana"